Как става преобразуването на слънчевата енергия. Принципът на преобразуване на слънчевата енергия в електричество Какво се случва в светлинната фаза на фотосинтезата
Прочетете също
Учебник за 10-11 клас
Глава III. Осигуряване на клетките с енергия
Всеки жив организъм, подобно на една клетка, е отворена система, т.е. обменя се с околен святматерия и енергия. Целият набор от ензимни реакции на метаболизма, протичащи в организма, се нарича метаболизъм (от гръцки "metabole" - трансформация). Метаболизмът се състои от взаимосвързани реакции на асимилация - синтез на макромолекулни съединения (протеини, нуклеинови киселини, полизахариди, липиди) и дисимилация - разделяне и окисляване на органични вещества, които вървят с преобразуването на енергия. Асимилацията, наричана още пластична обмяна, е невъзможна без енергията, освободена в резултат на дисимилацията (енергиен обмен). Дисимилацията от своя страна не се извършва без ензими, образувани в резултат на пластичния метаболизъм.
Всяка проява на жизнена дейност (поглъщане на вода и разтворени в нея неорганични съединения, синтез на органични вещества, разграждане на полимери в мономери, генериране на топлина, движение и т.н.) изисква разход на енергия.
Основният източник на енергия за всички живи същества, които обитават нашата планета, е енергията на слънчевата светлина. Само клетки от зелени растения, едноклетъчни водорасли, зелени и лилави бактерии обаче го използват директно. Тези клетки, благодарение на енергията на слънчевата светлина, са в състояние да синтезират органични вещества - въглехидрати, мазнини, протеини, нуклеинови киселини. Биосинтезата, която възниква при използване на светлинна енергия, се нарича фотосинтеза. Организмите, способни на фотосинтеза, се наричат фотоавтотрофи.
Изходните вещества за фотосинтезата са вода, въглероден диоксид от земната атмосфера, както и неорганични соли на азот, фосфор, сяра от водни тела и почва. Молекулите на атмосферния азот (N 2) също са източник на азот, който се усвоява от бактерии, живеещи в почвата и в кореновите възли, главно на бобови растения. В този случай газообразният азот преминава в състава на молекулата на амоняка - NH3, който впоследствие се използва за синтеза на аминокиселини, протеини, нуклеинови киселини и други азотсъдържащи съединения. Нодулните бактерии и бобовите растения се нуждаят едни от други. Съвместно взаимноизгодно съществуване различни видовеорганизмите се наричат симбиоза.
В допълнение към фотоавтотрофите, някои бактерии (водородни, нитрифициращи, серни бактерии и др.) Също така са способни да синтезират органични вещества от неорганични вещества. Те осъществяват този синтез благодарение на енергията, освободена при окисляването на неорганичните вещества. Те се наричат хемоавтотрофи. Процесът на хемосинтеза е открит през 1887 г. от руския микробиолог С. Н. Виноградски.
Всички живи същества на нашата планета, които не могат да синтезират органични вещества от неорганични съединения, се наричат хетеротрофи. Всички животни и хора живеят от енергията на Слънцето, съхранявана от растенията, преобразувана в енергията на химичните връзки на новосинтезирани органични съединения.
Трябва да се отбележи, че както фотосинтетичните, така и хемосинтетичните организми също могат да получават енергия поради окисляването на органични вещества. Хетеротрофите обаче получават тези вещества готови отвън, докато автотрофите ги синтезират от неорганични съединения.
Фотосинтетичните клетки абсорбират въглероден диоксид от атмосферата и отделят кислород в нея. Преди появата на фотосинтетичните клетки на нашата планета земната атмосфера е била лишена от кислород. С появата на фотосинтезиращите организми постепенното изпълване на атмосферата с кислород доведе до появата на клетки с нов тип енергиен апарат. Това са клетки, които произвеждат енергия чрез окисляване на готови органични съединения, главно въглехидрати и мазнини, с участието на атмосферния кислород като окислител. Когато органичните съединения се окисляват, се освобождава енергия.
В резултат на насищането на атмосферата с кислород възникват аеробни клетки, които могат да използват кислород за енергия.
§ 11. Фотосинтеза. Преобразуване на светлинна енергия в енергия на химична връзка
Първите клетки, способни да използват енергията на слънчевата светлина, са се появили на Земята преди около 4 милиарда години в архейската ера. Това бяха цианобактерии (от гръцки "cyanos" - синьо). Техните вкаменени останки са открити в шистови пластове, датиращи от този период от историята на Земята. Отне още 1,5 милиарда години за насищане на земната атмосфера с кислород и появата на аеробни клетки.
Очевидно е, че ролята на растенията и другите фотосинтезиращи организми в развитието и поддържането на живота на нашата планета е изключително голяма: те преобразуват енергията на слънчевата светлина в енергията на химичните връзки на органичните съединения, която след това се използва от всички останали живи организми. същества; те насищат атмосферата на Земята с кислород, който служи за окисляване на органичните вещества и по този начин извлича химическата енергия, съхранявана в тях от аеробните клетки; И накрая, някои растителни видове в симбиоза с азотфиксиращи бактерии въвеждат газообразен азот от атмосферата в молекулите на амоняка, неговите соли и органичните азотсъдържащи съединения.
Ролята на зелените растения в живота на планетата не може да бъде надценена. Запазването и разширяването на зелената покривка на Земята е от изключително значение за всички живи същества, населяващи нашата планета.
Съхранение на светлинна енергия в биологични "акумулатори".Потокът от слънчева светлина носи светлинни вълни с различна дължина. Растенията с помощта на светлинни "антени" (това са главно молекули на хлорофил) поглъщат светлинни вълни от червената и синята част на спектъра. Хлорофилът пропуска светлинните вълни от зелената част на спектъра без забавяне и затова растенията имат зелен цвят.
С помощта на светлинна енергия електрон в състава на молекулата на хлорофила се прехвърля на по-високо енергийно ниво. Освен това този високоенергиен електрон, сякаш на стъпала, скача по веригата от електронни носители, губейки енергия. В този случай енергията на електроните се изразходва за „зареждане“ на вид биологични „акумулатори“. Без да навлизаме в химичните особености на тяхната структура, нека кажем, че една от тях е аденозинтрифосфорната киселина, която се нарича още аденозинтрифосфат (накратко АТФ). Както вече беше споменато в § 6, АТФ съдържа три остатъка от фосфорна киселина, свързани един с друг, които са прикрепени към аденозин. Схематично АТФ може да се опише с формулата: аденозин-Р-Р~Р, където Р е остатък от фосфорна киселина. В химическата връзка между втория и третия краен фосфат се съхранява енергия, която електронът отдава (такава специална химическа връзка е изобразена с вълнообразна линия). Това се дължи на факта, че когато един електрон прехвърли енергията си на аденозин дифосфат (аденозин-FF, ADP), се добавя друг фосфат: ADP + F + E → ATP, където E е енергията на електрона, която се съхранява в ATP. Когато АТФ се разцепва от ензима аденозин трифосфатаза (АТФаза), крайният фосфат се отцепва и се освобождава енергия:
В растителната клетка енергията на АТФ се използва за транспортиране на вода и соли, за делене, растеж и движение на клетките (спомнете си как главата на слънчогледа се върти след Слънцето).
АТФ енергията е необходима за синтеза на глюкоза, нишесте, целулоза и други органични съединения в растенията. Но за синтеза на органични вещества в растенията е необходим още един биологичен „акумулатор“, който съхранява енергията на светлината. Тази батерия има непроизносимо дълго име: никотин амид аденин динуклеотид фосфат (накратко NADP, произнася се „nad-ef“). Това съединение съществува в редуцирана, високоенергийна форма: NADP-N (произнася се „над-еф-аш“).
Губещата енергия окислена форма на това съединение е NADP+ (произнася се „nad-ef-plus“). Губейки един водороден атом и един електрон, NADP-N се превръща в NADP + и редуцира въглеродния диоксид (с участието на водни молекули) до глюкоза C 6 H 12 0 6; липсващите протони (H +) се вземат от водната среда. В опростена форма този процес може да бъде написан като химическо уравнение:
Въпреки това, когато въглероден диоксид и вода се смесят, глюкоза не се образува. Това изисква не само възстановителната сила на NADPH, но и енергията на АТФ и съединение, което свързва CO 2, което се използва в междинните етапи на синтеза на глюкозата, както и редица ензими - биологични катализатори за този процес.
Фотолиза на вода.Как се произвежда кислород по време на фотосинтеза? Факт е, че енергията на светлината се изразходва и за разделянето на водната молекула - фотолиза. В този случай се образуват протони (Н +), електрони (О и свободен кислород:
Електроните, произведени по време на фотолиза, компенсират загубата им от хлорофила (както се казва, те запълват „дупката“, която е възникнала в хлорофила). Част от електроните, с участието на протони, редуцира NADP + до NADP-H. Кислородът е страничен продукт от тази реакция (фиг. 19). Както може да се види от обобщеното уравнение за синтеза на глюкоза, при този процес се отделя кислород.
Когато растенията използват енергията на слънчевата светлина, те не се нуждаят от кислород. Въпреки това, при липса на слънчева светлина, растенията стават аероби. В тъмнината на нощта те консумират кислород и окисляват глюкозата, фруктозата, нишестето и други съединения, складирани през деня, като животните в това.
Светли и тъмни фази на фотосинтезата.В процеса на фотосинтеза се разграничават светли и тъмни фази. Когато растенията са осветени, енергията на светлината се преобразува в енергията на химичните връзки на ATP и NADP-H. Енергията на тези съединения лесно се освобождава и използва вътре в растителната клетка за различни цели, предимно за синтеза на глюкоза и други органични съединения. Следователно този начален етап на фотосинтезата се нарича светлинна фаза. Без осветяване със слънчева или изкуствена светлина, в чийто спектър има червени и сини лъчи, синтезът на АТФ и NADP-H не се извършва в растителната клетка. Когато обаче ATP и NADP-H молекулите вече са натрупани в растителната клетка, синтезът на глюкоза може да се случи и на тъмно, без участието на светлина. За тези биохимични реакции не е необходимо осветление, тъй като те вече са снабдени със светлинна енергия, съхранявана в биологични "акумулатори". Този етап от фотосинтезата се нарича температурна фаза.

Ориз. 19. Схема на фотосинтезата
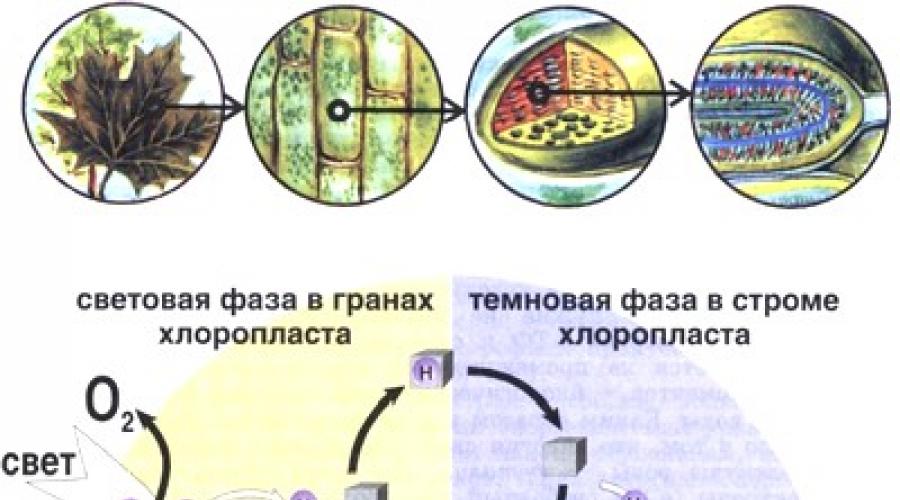
Всички реакции на фотосинтеза протичат в хлоропласти - удебелени овални или кръгли образувания, разположени в цитоплазмата на растителна клетка (хлоропластите вече бяха разгледани накратко в § 9). Всяка клетка съдържа 40-50 хлоропласта. Хлоропластите са ограничени отвън с двойна мембрана, а вътре в тях има тънки плоски торбички - тилакоиди, също ограничени от мембрани. Тилакоидите съдържат хлорофил, преносители на електрони и всички ензими, участващи в светлинната фаза на фотосинтезата, както и ADP, ATP, NADP + и NADP-H. Десетки тилакоиди са плътно опаковани в купчини, наречени грана. в вътрешно пространствомежду граната - в стромата на хлоропластите - има ензими, участващи в редукцията на CO 2 до глюкоза поради енергията на продуктите на светлинната фаза на фотосинтезата - ATP и NADP-H. Следователно реакциите на тъмната фаза на фотосинтезата се появяват в стромата, тясно свързана със светлата фаза, която се разгръща в тилакоидите. Светлата и тъмната фаза на фотосинтезата са показани схематично на фигура 19.
Хлоропластите имат собствен генетичен апарат - ДНК молекули и автономно се възпроизвеждат вътре в клетките. Смята се, че преди повече от 1,5 милиарда години те са били свободни микроорганизми, които са станали симбионти на растителни клетки.
- Обяснете защо казваме, че енергията за живот на Земята първоначално се доставя от Слънцето.
- Обяснете защо фотосинтезата използва въглероден диоксид и вода и какъв е източникът на страничния продукт на фотосинтезата, кислород.
- Как са свързани проблемите на фотосинтезата и снабдяването с храна на населението на Земята?
- Защо фотосинтезата преобразува енергията на слънчевата светлина, падаща върху листа, в енергия, съхранявана в органични съединения с ефективност само около 1%? Каква е съдбата на останалата енергия?
- Попълнете таблицата.
Историята на изучаването на фотосинтезата датира от август 1771 г., когато английският теолог, философ и любител натуралист Джоузеф Пристли (1733-1804) открива, че растенията могат да "коригират" свойствата на въздуха, който променя състава си в резултат на изгаряне или живот на животните. Пристли показа, че в присъствието на растения "разваленият" въздух отново става подходящ за изгаряне и поддържане живота на животните.
В хода на по-нататъшни изследвания от Ingenhaus, Senebier, Saussure, Bussengo и други учени беше установено, че растенията, когато са осветени, отделят кислород и абсорбират въглероден диоксид от въздуха. Растенията синтезират органични вещества от въглероден диоксид и вода. Този процес се нарича фотосинтеза.
Робърт Майер, който открива закона за запазване на енергията, предполага през 1845 г., че растенията преобразуват енергията на слънчевата светлина в енергията на химичните съединения, образувани по време на фотосинтезата. Според него "слънчевите лъчи, разпространяващи се в космоса, се "улавят" и съхраняват за по-нататъшно използване при необходимост". Впоследствие руският учен К.А. Тимирязев убедително доказва, че най-важната роля в използването на слънчевата енергия от растенията играят молекулите на хлорофила, присъстващи в зелените листа.
Въглехидратите (захарите), образувани по време на фотосинтезата, се използват като източник на енергия и строителен материал за синтеза на различни органични съединения в растенията и животните. При висшите растения процесите на фотосинтеза протичат в хлоропластите - специализирани енергопреобразуващи органели на растителна клетка.
Схематично представяне на хлоропласта е показано на фиг. един.
Под двойната обвивка на хлоропласта, състояща се от външна и вътрешна мембрана, има разширени мембранни структури, които образуват затворени везикули, наречени тилакоиди. Тилакоидните мембрани се състоят от два слоя липидни молекули, които включват макромолекулни фотосинтетични протеинови комплекси. В хлоропластите на висшите растения тилакоидите са групирани в grana, които са купчини от дисковидни, сплескани и плътно притиснати един към друг тилакоиди. Междузърнестите тилакоиди, излизащи от тях, са продължение на отделни тилакоиди на граната. Пространството между мембраната на хлоропласта и тилакоидите се нарича строма. Стромата съдържа РНК, ДНК, хлоропластни молекули, рибозоми, нишестени зърна и множество ензими, включително тези, които осигуряват усвояването на CO2 от растенията.
Изданието е реализирано с подкрепата на Sushi E'xpress. Компанията Sushi E'xpress предоставя услуги за доставка на суши в Новосибирск. Поръчвайки суши от Sushi E’xpress, бързо ще получите вкусно и здравословно ястие, приготвено от професионални готвачи с най-пресните продукти с най-високо качество. Посещавайки сайта на Sushi E’xpress можете да се запознаете с цените и състава на предлаганите рулца, което ще ви помогне да вземете решение при избора на ястие. За заявка за доставка на суши се обадете на тел. 239-55-87
Светли и тъмни етапи на фотосинтезата
Според модерни идеи, фотосинтезата е поредица от фотофизични и биохимични процеси, в резултат на които растенията синтезират въглехидрати (захари), използвайки енергията на слънчевата светлина. Многобройните етапи на фотосинтезата обикновено се разделят на две големи групи процеси - светли и тъмни фази.
Прието е светлинните етапи на фотосинтезата да се наричат набор от процеси, в резултат на които, благодарение на енергията на светлината, се синтезират молекули на аденозин трифосфат (АТФ) и се образува редуциран никотинамид аденин динуклеотид фосфат (NADPH), a съединение с висок редукционен потенциал. Молекулите на АТФ действат като универсален източник на енергия в клетката. Известно е, че енергията на високоенергийните (т.е. богати на енергия) фосфатни връзки на молекулата на АТФ се използва в повечето енергоемки биохимични процеси.
Светлинните процеси на фотосинтеза протичат в тилакоидите, чиито мембрани съдържат основните компоненти на фотосинтетичния апарат на растенията - пигментно-протеинови и електрон-транспортни комплекси, събиращи светлина, както и АТФ-синтазния комплекс, който катализира образуването на АТФ от аденозин дифосфат (ADP) и неорганичен фосфат (F i) (ADP + F i → ATP + H 2 O). Така, в резултат на светлинните етапи на фотосинтезата, енергията на светлината, погълната от растенията, се съхранява под формата на макроергични химични връзки на АТФ молекули и силния редуциращ агент NADP H, които се използват за синтез на въглехидрати в т.нар. тъмни етапи на фотосинтезата.
Тъмните етапи на фотосинтезата обикновено се наричат набор от биохимични реакции, които водят до усвояването на атмосферния въглероден диоксид (CO 2 ) от растенията и образуването на въглехидрати. Цикълът на тъмните биохимични трансформации, водещи до синтеза на органични съединения от CO2 и вода, се нарича цикъл на Калвин-Бенсън на името на авторите, които имат решаващ принос в изследването на тези процеси. За разлика от електронния транспорт и АТФ синтазните комплекси, които се намират в тилакоидната мембрана, ензимите, които катализират "тъмните" реакции на фотосинтезата, са разтворени в стромата. Когато хлоропластната мембрана се разруши, тези ензими се измиват от стромата, в резултат на което хлоропластите губят способността си да абсорбират въглероден диоксид.
В резултат на трансформации на редица органични съединения в цикъла на Калвин-Бенсън от три молекули CO 2 и вода в хлоропластите се образува молекула глицералдехид-3-фосфат, която има химична формула CHO–CHOH–CH 2 O– PO 3 2-. В същото време, три ATP молекули и две NADP H молекули се консумират на една CO 2 молекула, включена в глицералдехид-3-фосфат.
За синтеза на органични съединения в цикъла на Калвин-Бенсън се използва енергията, освободена по време на хидролизата на макроергичните фосфатни връзки на ATP молекулите (реакция ATP + H 2 O → ADP + F i) и силния редукционен потенциал на NADP H молекулите , Основната част от молекулите, образувани в хлоропласта Глицералдехид-3-фосфат навлиза в цитозола на растителната клетка, където се превръща във фруктозо-6-фосфат и глюкозо-6-фосфат, които в хода на по-нататъшни трансформации, образуват захарофосфат, прекурсор на захарозата. От молекулите на глицералдехид-3-фосфат, останали в хлоропласта, се синтезира нишесте.
Преобразуване на енергия във фотосинтетични реакционни центрове
Фотосинтетичните енергопреобразуващи комплекси от растения, водорасли и фотосинтезиращи бактерии са добре проучени. Инсталиран химичен състави пространствената структура на енерготрансформиращите протеинови комплекси е изяснена последователността на процесите на енергийна трансформация. Въпреки разликите в състава и молекулярната структура на фотосинтетичния апарат, съществуват общи закономерности в процесите на преобразуване на енергия във фотореакционните центрове на всички фотосинтетични организми. Във фотосинтетичните системи както от растителен, така и от бактериален произход, една структурна и функционална връзка на фотосинтетичния апарат е фотосистема, който включва антена за събиране на светлина, фотохимичен реакционен център и свързани с него молекули - носители на електрони.
Помислете първо основни принципипреобразуване на енергията на слънчевата светлина, характерно за всички фотосинтетични системи, след което ще се спрем по-подробно на примера за функционирането на фотореакционните центрове и веригата за пренос на електрони на хлоропластите във висшите растения.
Антена за събиране на светлина (поглъщане на светлина, миграция на енергия към реакционния център)
Първият елементарен акт на фотосинтезата е поглъщането на светлина от хлорофилни молекули или спомагателни пигменти, които са част от специален комплекс пигмент-протеин, наречен антена за събиране на светлина. Антената за събиране на светлина е макромолекулен комплекс, предназначен да улавя ефективно светлината. В хлоропластите антенният комплекс съдържа голям брой (до няколкостотин) хлорофилни молекули и известно количество спомагателни пигменти (каротеноиди), силно свързани с протеина.
При ярка слънчева светлина една молекула на хлорофила поглъща светлинни кванти относително рядко, средно не повече от 10 пъти в секунда. Въпреки това, тъй като един фотореакционен център представлява голям брой хлорофилни молекули (200–400), дори при относително нисък интензитет на светлината, падаща върху лист при условия на засенчване на растението, реакционният център се активира доста често. Ансамбълът от светлопоглъщащи пигменти всъщност играе ролята на антена, която поради доста големия си размер ефективно улавя слънчевата светлина и насочва енергията й към реакционния център. Сенколюбивите растения обикновено имат по-голям размерантена за събиране на светлина в сравнение с растенията, растящи при условия на силна светлина.
Молекулите на хлорофила са основните пигменти за събиране на светлина в растенията. аи хлорофил bабсорбиране на видима светлина с дължина на вълната λ ≤ 700–730 nm. Изолираните хлорофилни молекули абсорбират светлина само в две сравнително тесни ленти на слънчевия спектър: при дължини на вълните 660–680 nm (червена светлина) и 430–450 nm (синьо-виолетова светлина), което, разбира се, ограничава ефективността на използването на целият спектър от слънчева светлина пада върху зелено листо.
Обаче спектралният състав на светлината, абсорбирана от антената за събиране на светлина, всъщност е много по-широк. Това се обяснява с факта, че спектърът на поглъщане на агрегирани форми на хлорофил, които са част от антената за събиране на светлина, е изместен към по-дълги дължини на вълните. Заедно с хлорофила, антената за събиране на светлина включва помощни пигменти, които повишават нейната ефективност поради факта, че те абсорбират светлина в тези спектрални области, в които молекулите на хлорофила, основният пигмент на антената за събиране на светлина, абсорбират светлината относително слабо.
В растенията спомагателните пигменти са каротеноиди, които абсорбират светлина в диапазона на дължината на вълната λ ≈ 450–480 nm; в клетките на фотосинтезиращите водорасли това са червени и сини пигменти: фикоеритрини в червени водорасли (λ ≈ 495–565 nm) и фикоцианини в синьо-зелени водорасли (λ ≈ 550–615 nm).
Поглъщането на светлинен квант от молекула на хлорофил (Chl) или спомагателен пигмент води до неговото възбуждане (електронът преминава на по-високо енергийно ниво):
Chl + hν → Chl*.
Енергията на възбудената хлорофилна молекула Chl * се прехвърля към молекулите на съседните пигменти, които от своя страна могат да я прехвърлят към други молекули на антената за събиране на светлина:
Chl* + Chl → Chl + Chl*.
 По този начин енергията на възбуждане може да мигрира през пигментната матрица, докато възбуждането в крайна сметка достигне фотореакционния център P (схематично представяне на този процес е показано на фиг. 2):
По този начин енергията на възбуждане може да мигрира през пигментната матрица, докато възбуждането в крайна сметка достигне фотореакционния център P (схематично представяне на този процес е показано на фиг. 2):
Chl* + P → Chl + P*.
Имайте предвид, че продължителността на съществуване на молекулите на хлорофила и други пигменти във възбудено състояние е много кратка, τ ≈ 10–10–10–9 s. Следователно има известна вероятност, че по пътя към реакционния център P енергията на такива краткотрайни възбудени състояния на пигменти може да бъде безполезно загубена - разсеяна в топлина или освободена под формата на светлинен квант (флуоресцентен феномен). В действителност обаче ефективността на миграцията на енергия към фотосинтетичния реакционен център е много висока. В случай, че реакционният център е в активно състояние, вероятността от загуба на енергия е, като правило, не повече от 10–15%. Такава висока ефективност на използване на енергията на слънчевата светлина се дължи на факта, че антената за събиране на светлина е високо подредена структура, която осигурява много добро взаимодействие на пигментите един с друг. Благодарение на това се постига висока скорост на пренос на енергия на възбуждане от молекули, които абсорбират светлина, към фотореакционния център. Средното време на "скок" на енергията на възбуждане от един пигмент към друг, като правило, е τ ≈ 10–12–10–11 s. Общото време на миграция на възбуждане към реакционния център обикновено не надвишава 10–10–10–9 s.
Фотохимичен реакционен център (трансфер на електрони, стабилизиране на отделените заряди)
Съвременните идеи за структурата на реакционния център и механизмите на първичните етапи на фотосинтезата са предшествани от трудовете на A.A. Красновски, който откри, че в присъствието на донори и акцептори на електрони, молекулите на хлорофила, възбудени от светлина, могат да бъдат обратимо редуцирани (приемане на електрон) и окисляване (даряване на електрон). Впоследствие Кок, Вит и Дуизенс откриват в растенията, водораслите и фотосинтезиращите бактерии специални пигменти от хлорофилна природа, наречени реакционни центрове, които се окисляват под действието на светлината и всъщност са първичните донори на електрони по време на фотосинтезата.
Фотохимичният реакционен център P е специална двойка (димер) от хлорофилни молекули, които действат като капан за енергията на възбуждане, блуждаеща през пигментната матрица на антената за събиране на светлина (фиг. 2). Точно както течността тече от стените на широка фуния към нейното тясно гърло, енергията на светлината, погълната от всички пигменти на антената за събиране на светлина, е насочена към реакционния център. Възбуждането на реакционния център инициира верига от по-нататъшни трансформации на светлинна енергия по време на фотосинтезата.
Последователността на процесите, протичащи след възбуждането на реакционния център P, и диаграмата на съответните промени в енергията на фотосистемата са показани схематично на фиг. 3.
Заедно с димера на хлорофил P, фотосинтетичният комплекс включва молекули на първични и вторични акцептори на електрони, които условно ще обозначим като A и B, както и първичния донор на електрони, молекулата D. към съседния му първичен акцептор на електрони A:
D(P*A)B → D(P + A –)B.
Така в резултат на много бърз (t ≈ 10–12 s) трансфер на електрони от P* към A се осъществява втората принципно важна стъпка в преобразуването на слънчевата енергия по време на фотосинтезата – разделяне на заряда в реакционния център. В този случай се образува силен редуциращ агент А - (донор на електрони) и силен окислител Р + (акцептор на електрони).
Молекулите P + и A - са разположени в мембраната асиметрично: в хлоропластите реакционният център P + е по-близо до повърхността на мембраната, обърната вътре в тилакоида, а акцепторът A - е разположен по-близо до външната страна. Следователно, в резултат на фотоиндуцираното разделяне на заряда, на мембраната възниква разлика в електрическите потенциали. Индуцираното от светлина разделяне на зарядите в реакционния център е подобно на генерирането на електрическа потенциална разлика в конвенционална фотоклетка. Все пак трябва да се подчертае, че за разлика от всички известни и широко използвани енергийни фотопреобразуватели в техниката, ефективността на работа на фотосинтетичните реакционни центрове е много висока. Ефективността на разделяне на заряда в активните фотосинтетични реакционни центрове като правило надвишава 90–95% (за най-добрите образци фотоклетки ефективността не надвишава 30%).
Какви механизми осигуряват такава висока ефективност на преобразуване на енергия в реакционните центрове? Защо електронът, прехвърлен към акцептора A, не се връща обратно към положително заредения окислен център P +? Стабилизирането на отделените заряди се осигурява главно поради вторичните процеси на електронен транспорт след прехвърлянето на електрон от P* към A. От редуцирания първичен акцептор A, електрон много бързо (за 10–10–10–9 s ) отива към вторичния акцептор на електрони B:
D(P + A –)B → D(P + A)B – .
В този случай се случва не само отстраняването на електрон от положително заредения реакционен център P +, но и енергията на цялата система също значително намалява (фиг. 3). Това означава, че за да прехвърли електрон в обратна посока (преход B - → A), той ще трябва да преодолее достатъчно висока енергийна бариера ΔE ≈ 0,3–0,4 eV, където ΔE е разликата в енергийните нива за двете състояния на системата, в която електронът е съответно на носителя А или В. Поради тази причина, за да се върне електронът обратно, от редуцираната молекула B към окислената молекула A, ще отнеме много повече време, отколкото за директния преход A - → B. С други думи, в посока напред електронът се пренася много по-бързо, отколкото обратното. Следователно, след прехвърлянето на електрон към вторичния акцептор B, вероятността за връщането му обратно и рекомбинацията с положително заредената "дупка" P + значително намалява.
Вторият фактор, допринасящ за стабилизирането на отделените заряди, е бързото неутрализиране на окисления фотореакционен център P + поради идването на електрона към P + от донора на електрон D:
D(P + A)B – → D + (PA)B – .
След като е получил електрон от донорната молекула D и се е върнал в първоначалното си редуцирано състояние P, реакционният център вече няма да може да приеме електрон от редуцираните акцептори, но сега е готов да се задейства повторно - да дари електрон на разположен до него окислен първичен акцептор А. Това е последователността от събития, които се случват във фотореакционните центрове на всички фотосинтетични системи.
Хлоропластна електротранспортна верига
В хлоропластите на висшите растения има две фотосистеми: фотосистема 1 (PS1) и фотосистема 2 (PS2), които се различават по състава на протеини, пигменти и оптични свойства. Антената за събиране на светлина PS1 абсорбира светлина с дължина на вълната λ ≤ 700–730 nm, а PS2 абсорбира светлина с дължина на вълната λ ≤ 680–700 nm. Индуцираното от светлина окисление на реакционните центрове PS1 и PS2 е придружено от тяхното обезцветяване, което се характеризира с промени в техните абсорбционни спектри при λ ≈ 700 и 680 nm. В съответствие с техните оптични характеристики, реакционните центрове PS1 и PS2 бяха наречени P 700 и P 680.
Двете фотосистеми са свързани помежду си чрез верига от електронни носители (фиг. 4). PS2 е източникът на електрони за PS1. Светлинно инициираното разделяне на заряда във фотореакционните центрове P 700 и P 680 осигурява трансфер на електрони от вода, разложена в PS2, до крайния акцептор на електрони, молекулата NADP+. Електрон-транспортната верига (ETC), свързваща две фотосистеми, включва молекули пластохинон, отделен електронен транспортен протеинов комплекс (така наречения b/f комплекс) и водоразтворимия протеин пластоцианин (Pc) като носители на електрони. На фиг. четири.
В PS2 един електрон се прехвърля от възбудения център P * 680 първо към първичния акцептор феофетин (Phe), а след това към молекулата на пластохинона Q A, здраво свързана с един от PS2 протеините,
Y(P* 680 Phe)Q A Q B → Y(P + 680 Phe –)Q A Q B → Y(P + 680 Phe)Q A – Q B .
След това електронът се прехвърля към втората молекула пластохинон Q B и P 680 получава електрон от първичния донор на електрони Y:
Y(P + 680 Phe)Q A – Q B → Y + (P 680 Phe)Q A Q B – .
 Молекулата на пластохинона, чиято химична формула и местоположение в липидната двуслойна мембрана е показано на фиг. 5 е в състояние да приеме два електрона. След като реакционният център PS2 се задейства два пъти, молекулата на пластохинона Q B ще получи два електрона:
Молекулата на пластохинона, чиято химична формула и местоположение в липидната двуслойна мембрана е показано на фиг. 5 е в състояние да приеме два електрона. След като реакционният център PS2 се задейства два пъти, молекулата на пластохинона Q B ще получи два електрона:
Q B + 2е – → Q B 2– .
Отрицателно заредената Q B 2– молекула има висок афинитет към водородните йони, които улавя от стромалното пространство. След протониране на редуцирания пластохинон Q B 2– (Q B 2– + 2H + → QH 2) се образува електрически неутрална форма на тази QH 2 молекула, която се нарича пластохинол (фиг. 5). Пластохинолът действа като мобилен носител на два електрона и два протона: след като напусне PS2, молекулата QH2 може лесно да се движи вътре в тилакоидната мембрана, осигурявайки свързване на PS2 с други транспортни комплекси за електрони.
Окисленият реакционен център PS2 P 680 има изключително висок електронен афинитет; е много силен окислител. Поради това водата, химически стабилно съединение, се разлага в PS2. Комплексът за разделяне на водата (WRC), включен в PS2, съдържа в активния си център група манганови йони (Mn 2+), които служат като донори на електрони за P 680 . Дарявайки електрони на окисления реакционен център, мангановите йони стават „акумулатори“ на положителни заряди, които участват пряко в реакцията на окисляване на водата. В резултат на последователно четирикратно активиране на реакционния център P 680, четири силни окислителни еквивалента (или четири „дупки“) се натрупват в Mn-съдържащия активен център на WRC под формата на окислени манганови йони (Mn 4+) , които, взаимодействайки с две водни молекули, катализират реакционната вода на разлагане:
2Mn 4+ + 2H 2 O → 2Mn 2+ + 4H + + O 2 .
По този начин, след последователното прехвърляне на четири електрона от WRC към P 680, настъпва синхронно разлагане на две водни молекули наведнъж, придружено от освобождаване на една кислородна молекула и четири водородни йона, които навлизат в интратилакоидното пространство на хлоропласта.
Молекулата на пластохинола QH2, образувана по време на функционирането на PS2, дифундира в липидния двоен слой на тилакоидната мембрана до b/f комплекса (фиг. 4 и 5). При сблъсък с b/f комплекса, молекулата QH 2 се свързва с него и след това му прехвърля два електрона. В този случай, за всяка молекула пластохинол, окислена от b/f комплекса, два водородни йона се освобождават вътре в тилакоида. На свой ред b/f комплексът служи като донор на електрони за пластоцианин (Pc), сравнително малък водоразтворим протеин, чийто активен център съдържа меден йон (реакциите на редукция и окисление на пластоцианин са придружени от промени във валентността на медта йон Cu 2+ + e – ↔Cu+). Пластоцианинът действа като връзка между b/f комплекса и PS1. Молекулата на пластоцианина се движи бързо в тилакоида, осигурявайки трансфер на електрон от b/f комплекса към PS1. От редуцирания пластоцианин, електронът отива директно в окислените реакционни центрове на PS1 – P 700 + (виж фиг. 4). По този начин, в резултат на съвместното действие на PS1 и PS2, два електрона от водната молекула, разложена в PS2, в крайна сметка се прехвърлят през електронната транспортна верига към NADP + молекулата, осигурявайки образуването на силен редуциращ агент NADP H.
Защо хлоропластите се нуждаят от две фотосистеми? Известно е, че фотосинтетичните бактерии, които използват различни органични и неорганични съединения (например H 2 S) като донор на електрони за намаляване на окислените реакционни центрове, успешно функционират с една фотосистема. Появата на две фотосистеми най-вероятно се дължи на факта, че енергията на един квант видима светлина не е достатъчна, за да осигури разлагането на водата и ефективното преминаване на електрона по целия път по веригата на молекулите носители от вода до NADP + . Преди около 3 милиарда години на Земята се появиха синьо-зелени водорасли или цианобактерии, които придобиха способността да използват водата като източник на електрони за намаляване на въглеродния диоксид. Сега се смята, че PS1 произлиза от зелени бактерии, а PS2 от лилави бактерии. След като PS2 се „съедини“ в една верига за пренос на електрони заедно с PS1 по време на еволюционния процес, стана възможно да се реши енергиен проблем- за преодоляване на доста голяма разлика в редокс потенциалите на двойките кислород / вода и NADP + / NADP H. Появата на фотосинтезиращи организми, способни да окисляват вода, се превърна в един от крайъгълни камъниразвитието на дивата природа на Земята. Първо, водораслите и зелените растения, след като са се "научили" да окисляват водата, са усвоили неизчерпаем източник на електрони за редуциране на NADP +. Второ, чрез разграждането на водата те изпълват земната атмосфера с молекулярен кислород, създавайки условия за бързо еволюционно развитие на организми, чиято енергия е свързана с аеробно дишане.
Свързване на процесите на транспорт на електрони с трансфер на протони и синтез на АТФ в хлоропластите
Прехвърлянето на електрон по CET по правило е придружено от намаляване на енергията. Този процес може да се оприличи на спонтанно движение на тяло по наклонена равнина. Намаляването на енергийното ниво на електрона по време на движението му по CET изобщо не означава, че прехвърлянето на електрона винаги е енергийно безполезен процес. При нормални условия за функциониране на хлоропластите по-голямата част от енергията, освободена по време на преноса на електрони, не отива на вятъра, а се използва за работата на специален енергопреобразуващ комплекс, наречен АТФ синтаза. Този комплекс катализира енергийно неблагоприятния процес на образуване на АТФ от ADP и неорганичен фосфат F i (реакция ADP + F i → ATP + H 2 O). В тази връзка е обичайно да се казва, че енергоотдаващите процеси на електронен транспорт са свързани с енергийно приемащи процеси на синтез на АТФ.
Най-важната роля в осигуряването на енергийно свързване в тилакоидните мембрани, както и във всички други енергопреобразуващи органели (митохондрии, хроматофори на фотосинтезиращи бактерии), играят процесите на протонен транспорт. Синтезът на АТФ е тясно свързан с преноса на три протона чрез АТФ синтаза от тилакоидите (3Н в +) към стромата (3Н навън +):
ADP + F i + 3H in + → ATP + H 2 O + 3H out +.
Този процес става възможен, тъй като поради асиметричното разположение на носителите в мембраната, функционирането на хлоропластите ETC води до натрупване на излишно количество протони вътре в тилакоида: водородните йони се абсорбират отвън на етапите на NADP + редукция и образуване на пластохинол и се освобождават вътре в тилакоидите на етапите на разлагане на водата и окисление на пластохинол (фиг. 4). Осветяването на хлоропластите води до значително (100-1000 пъти) увеличение на концентрацията на водородни йони вътре в тилакоидите.
И така, разгледахме верига от събития, по време на които енергията на слънчевата светлина се съхранява под формата на енергия от високоенергийни химични съединения - ATP и NADP H. Тези продукти от светлинния етап на фотосинтезата се използват в тъмните етапи за образуване органични съединения (въглехидрати) от въглероден диоксид и вода. Основните етапи на преобразуване на енергията, водещи до образуването на ATP и NADP H, включват следните процеси: 1) абсорбция на светлинна енергия от пигментите на антената за събиране на светлина; 2) прехвърляне на енергия на възбуждане към фотореакционния център; 3) окисляване на фотореакционния център и стабилизиране на отделените заряди; 4) пренос на електрони по електротранспортната верига, образуване на NADP H; 5) трансмембранен пренос на водородни йони; 6) Синтез на АТФ.
1. Албъртс Б., Брей Д., Луис Дж., Робъртс К., Уотсън Дж.Молекулярна биология на клетката. Т. 1. - М .: Мир, 1994. 2-ро изд.
2. Кукушкин А.К., Тихонов А.Н.Лекции по биофизика на фотосинтезата на растенията. – М.: Издателство на Московския държавен университет, 1988 г.
3. Никълс Д.Д.Биоенергетика. Въведение в хемиосмотичната теория. – М.: Мир, 1985.
4. Скулачев В.П.Енергия на биологичните мембрани. – М.: Наука, 1989.
Как енергията на слънчевата светлина в светлата и тъмната фаза на фотосинтезата се превръща в енергията на химичните връзки на глюкозата? Обяснете отговора.
Отговор
В светлинната фаза на фотосинтезата енергията на слънчевата светлина се преобразува в енергията на възбудените електрони, а след това енергията на възбудените електрони се преобразува в енергията на АТФ и NADP-H2. В тъмната фаза на фотосинтезата енергията на АТФ и NADP-H2 се преобразува в енергията на глюкозните химични връзки.
Какво се случва по време на светлинната фаза на фотосинтезата?
Отговор
Електроните на хлорофила, възбудени от енергията на светлината, преминават по електротранспортните вериги, тяхната енергия се съхранява в ATP и NADP-H2. Настъпва фотолиза на водата, освобождава се кислород.
Кои са основните процеси, протичащи по време на тъмната фаза на фотосинтезата?
Отговор
От въглероден диоксид, получен от атмосферата, и водород, получен в светлинната фаза, се образува глюкоза поради енергията на АТФ, получена в светлинната фаза.
Каква е функцията на хлорофила в растителната клетка?
Отговор
Хлорофилът участва в процеса на фотосинтеза: в светлинната фаза хлорофилът абсорбира светлина, хлорофилният електрон получава светлинна енергия, откъсва се и преминава по веригата за транспортиране на електрони.
Каква роля играят електроните на хлорофила във фотосинтезата?
Отговор
Електроните на хлорофила, възбудени от слънчевата светлина, преминават през електронни транспортни вериги и предават енергията си за образуването на ATP и NADP-H2.
На какъв етап от фотосинтезата се произвежда свободен кислород?
Отговор
В светлинната фаза, по време на фотолизата на водата.
По време на коя фаза на фотосинтезата се извършва синтеза на АТФ?
Отговор
светлинна фаза.
Какъв е източникът на кислород по време на фотосинтеза?
Отговор
Вода (по време на фотолизата на водата се отделя кислород).
Скоростта на фотосинтезата зависи от ограничаващи (ограничаващи) фактори, сред които светлина, концентрация на въглероден диоксид, температура. Защо тези фактори са ограничаващи реакциите на фотосинтезата?
Отговор
Светлината е необходима за възбуждането на хлорофила, тя доставя енергия за процеса на фотосинтеза. Въглеродният диоксид е необходим в тъмната фаза на фотосинтезата; от него се синтезира глюкоза. Промяната в температурата води до денатурация на ензимите, реакциите на фотосинтезата се забавят.
При какви метаболитни реакции в растенията въглеродният диоксид е изходното вещество за синтеза на въглехидрати?
Отговор
в реакциите на фотосинтезата.
В листата на растенията процесът на фотосинтеза протича интензивно. Среща ли се в зрели и неузрели плодове? Обяснете отговора.
Отговор
Фотосинтезата се извършва в зелените части на растенията, изложени на светлина. По този начин фотосинтезата се извършва в кожата на зелените плодове. Вътре в плода и в кожата на зрели (не зелени) плодове не се извършва фотосинтеза.
Много от нас са се сблъсквали със слънчеви клетки по един или друг начин. Някой е използвал или използва слънчеви панели за генериране на електричество за битови нужди, някой използва малък слънчев панел, за да зарежда любимата си джаджа у дома. полеви условия, и някой със сигурност е видял малка слънчева клетка на микрокалкулатор. Някои дори имаха късмета да ги посетят.
Но замисляли ли сте се как протича процесът на преобразуване на слънчевата енергия в електрическа? Какво физическо явление е в основата на работата на всички тези слънчеви клетки? Нека се обърнем към физиката и да разберем процеса на генериране в детайли.
От самото начало е очевидно, че източникът на енергия тук е слънчевата светлина или, казано научен, тя се получава от фотони на слънчева радиация. Тези фотони могат да се представят като поток от елементарни частици, непрекъснато движещи се от Слънцето, всяка от които има енергия и следователно целият светлинен поток носи някакъв вид енергия.
От всеки квадратен метър от повърхността на Слънцето непрекъснато се излъчва 63 MW енергия под формата на радиация! Максималният интензитет на това лъчение попада в обхвата на видимия спектър -.
И така, учените са установили, че енергийната плътност на потока слънчева светлина на разстояние от Слънцето до Земята от 149 600 000 километра, след преминаване през атмосферата и при достигане на повърхността на нашата планета, е средно приблизително 900 вата на квадратен метър.
Тук можете да вземете тази енергия и да се опитате да получите електричество от нея, тоест да преобразувате енергията на слънчевия светлинен поток в енергията на движещи се заредени частици, с други думи, в.

За да преобразуваме светлината в електричество, имаме нужда от фотоволтаичен преобразувател. Такива преобразуватели са много разпространени, те се намират в свободна продажба, това са така наречените слънчеви клетки - фотоелектрически преобразуватели под формата на пластини, изрязани от силиций.
Най-добрите са монокристални, те имат ефективност около 18%, тоест, ако фотонният поток от слънцето има енергийна плътност 900 W / кв. м, тогава можете да разчитате на получаване на 160 W електроенергия на квадратен метър на батерия, сглобена от такива клетки.
Тук действа феномен, наречен "фотоелектричен ефект". Фотоелектричен ефект или фотоелектричен ефект- това е явлението на излъчване на електрони от вещество (явлението на изтегляне на електрони от атомите на вещество) под въздействието на светлина или друго електромагнитно излъчване.
Още през 1900 г. Макс Планк, бащата на квантовата физика, предполага, че светлината се излъчва и абсорбира на отделни порции или кванти, които по-късно, а именно през 1926 г., химикът Гилбърт Луис ще нарече "фотони".

Всеки фотон има енергия, която може да се определи по формулата E = hv – константата на Планк по честотата на излъчване.
В съответствие с идеята на Макс Планк, феноменът, открит през 1887 г. от Херц и след това задълбочено изследван от 1888 до 1890 г. от Столетов, стана обясним. Александър Столетов експериментално изследва фотоелектричния ефект и установи три закона на фотоелектричния ефект (закони на Столетов):
При постоянен спектрален състав на електромагнитното лъчение, падащо върху фотокатода, фототокът на насищане е пропорционален на енергийното осветяване на катода (в противен случай: броят на фотоелектроните, избити от катода за 1 s, е право пропорционален на интензитета на излъчване).
Максималната начална скорост на фотоелектроните не зависи от интензитета на падащата светлина, а се определя само от нейната честота.
За всяко вещество има червена граница на фотоелектричния ефект, тоест минималната честота на светлината (в зависимост от химическата природа на веществото и състоянието на повърхността), под която фотоелектричният ефект е невъзможен.
По-късно, през 1905 г., Айнщайн ще изясни теорията за фотоелектричния ефект. Той ще покаже как квантовата теория на светлината и законът за запазване и трансформация на енергията обясняват идеално какво се случва и какво се наблюдава. Айнщайн записва уравнението за фотоелектричния ефект, за което получава Нобелова награда през 1921 г.:

Работната функция И тук е минималната работа, която един електрон трябва да извърши, за да напусне атом на вещество. Вторият мандат - кинетична енергияелектрон след излизане.
Тоест фотонът се поглъща от електрона на атома, поради което кинетичната енергия на електрона в атома се увеличава с количеството енергия на погълнатия фотон.
Част от тази енергия се изразходва за излизане на електрона от атома, електронът напуска атома и получава възможност да се движи свободно. А насочено движещите се електрони не са нищо друго освен електрически ток или фототок. В резултат на това можем да говорим за появата на ЕМП в дадено вещество в резултат на фотоелектричния ефект.

Това е, Соларната батерия работи благодарение на действащия в нея фотоелектричен ефект.Но къде отиват "избитите" електрони във фотоелектрическия преобразувател? Фотоелектричен преобразувател или слънчева клетка или фотоклетка е, следователно, фотоелектрическият ефект в него се появява необичайно, това е вътрешен фотоелектричен ефект и дори има специално име "фотоелектричен ефект на вентила".
Под влияние на слънчевата светлина p-n преходВърху полупроводника се появява фотоелектричен ефект и се появява ЕМП, но електроните не напускат фотоклетката, всичко се случва в блокиращия слой, когато електроните напускат една част на тялото, премествайки се в друга част от него.
Силицият в земната кора е 30% от нейната маса, така че се използва навсякъде. Характеристика на полупроводниците като цяло е, че те не са нито проводници, нито диелектрици, тяхната проводимост зависи от концентрацията на примеси, от температурата и от излагането на радиация.
Забранената зона в полупроводника е няколко електронволта и това е само енергийната разлика между горното ниво на валентната лента на атомите, от която излизат електроните, и долното ниво на проводимата зона. Силицият има забранена лента от 1,12 eV - точно това, което е необходимо за поглъщане на слънчевата радиация.

И така, p-n преход. Легираните силициеви слоеве във фотоклетката образуват p-n преход. Тук се получава енергийна бариера за електроните, те напускат валентната зона и се движат само в една посока, дупките се движат в обратна посока. Така се получава токът в соларната клетка, тоест става генерирането на електричество от слънчева светлина.
P-n преходът, изложен на фотони, не позволява на носителите на заряд - електрони и дупки - да се движат в друга посока, освен само в една посока, те се разделят и завършват от противоположните страни на бариерата. И като бъде свързан към веригата на натоварване чрез горния и долния електрод, фотоволтаичният преобразувател, изложен на слънчева светлина, ще създаде във външната верига.
резюмедруги презентации"Въпроси на Единния държавен изпит по биология 2013" - Генотипът на организма. симбиотична връзка. протеинова молекула. Основни грешки. Тютюнев дим. Прилики и разлики между мутационната и комбинираната изменчивост. Нуклеотид. Колко клетки се образуват в резултат на мейозата. Протеинови молекули. Кръвна група. Синдром на Даун. Триплет от нуклеотиди. Етапи на енергийния метаболизъм. Консултация по биология. тромбоцити. Променливост на комбинацията. Способността за химио-автотрофно хранене.
„Болести на отделителната система” – Цистит. Уретрит. диабетна нифропатия. Нефрогенна анемия. Уролитиаза заболяване. пиелонефрит. Хидронефроза. Поликистоза на бъбреците. Бъбречна колика. Амилоидоза на бъбреците. Остри заболявания на отделителната система. простатит.
"Палеогенски период" - Климат. Климатът дори беше тропически. кайнозойска ера. Костни риби. Олигоцен. Началото на палеогена. Животински свят. Диатрима. палеоген. Ветрилоопашати беззъби птици. Горен еоцен. Палеогенски период.
„Въпроси на Единния държавен изпит по биология“ - Разцветът на класа Bird. Какъв вид размножаване е безполов. Каква структура на хлоропласта съдържа ензими. Установете съответствие между знака и органа. соматични мутации. външен вид на тъканите. Възприемане на раздразнение. При гръбначните животни органът на слуха се е променил в процеса на еволюция. Анализ на резултатите от изпита по биология. Плацентация. Появата на белите дробове. Структурата на окото. Колко автозоми има в ядрата на соматичните клетки.
"Правила за здравословно хранене" - Организация на столове с пълен цикъл. Изпълнение на програмата. Пиле. Резултати от изпълнението на програмата. Протеин. Правилното хранене е начин на живот. Здравословна храна. Правилното хранене. Теории за храненето. Храна в училище. Класическата теория за балансираното хранене. Цялостна реорганизация на системата за училищно хранене. Диети. Цели и задачи на програмата. Има 2 варианта за разработване на план за хранене в училище.
„Производство на млечни продукти” – Изследване качеството на млякото. Строителство. животновъдна индустрия. Доклад от обиколката. Определяне на въглехидрати в мляко. Старицки завод за масло и сирене. Изречение. Трудове на критици и анализатори. Произведено мляко. Приносът на великия учен. Приносът на Дмитрий Иванович Менделеев за развитието на млечната промишленост. Истински учен. Поръчка. Развитие на сирене. Идеи. Определение за мазнини. Имоти съставни частимляко.