Jak probíhá přeměna sluneční energie. Princip přeměny sluneční energie na elektřinu Co se děje ve světelné fázi fotosyntézy
Přečtěte si také
Učebnice pro ročníky 10-11
Kapitola III. Zásobování buněk energií
Jakýkoli živý organismus, stejně jako jedna buňka, je otevřený systém, to znamená, že si vyměňuje hmotu a energii s prostředím. Celý soubor enzymatických reakcí metabolismu probíhající v těle se nazývá metabolismus (z řeckého „metabole“ – přeměna). Metabolismus se skládá ze vzájemně souvisejících reakcí asimilace - syntézy makromolekulárních sloučenin (proteinů, nukleových kyselin, polysacharidů, lipidů) a disimilace - štěpení a oxidace organických látek, které jdou s přeměnou energie. Asimilace, nazývaná také plastická výměna, je nemožná bez energie uvolněné v důsledku disimilace (energetické výměny). Disimilace se zase neobejde bez enzymů vzniklých v důsledku metabolismu plastů.
Jakýkoli projev vitální činnosti (absorpce vody a v ní rozpuštěných anorganických sloučenin, syntéza organických látek, štěpení polymerů na monomery, tvorba tepla, pohyb atd.) vyžaduje vynaložení energie.
Hlavním zdrojem energie pro všechny živé bytosti, které obývají naši planetu, je energie slunečního světla. Přímo jej však využívají pouze buňky zelených rostlin, jednobuněčných řas, zelených a fialových bakterií. Tyto buňky jsou díky energii slunečního záření schopny syntetizovat organické látky – sacharidy, tuky, bílkoviny, nukleové kyseliny. Biosyntéza, ke které dochází při použití světelné energie, se nazývá fotosyntéza. Organismy schopné fotosyntézy se nazývají fotoautotrofy.
Výchozími látkami pro fotosyntézu jsou voda, oxid uhličitý zemské atmosféry, jakož i anorganické soli dusíku, fosforu, síry z vodních ploch a půdy. Zdrojem dusíku jsou také molekuly vzdušného dusíku (N 2), které jsou asimilovány bakteriemi žijícími v půdě a v kořenových uzlinách především luskovin. V tomto případě plynný dusík přechází do složení molekuly amoniaku - NH 3, který je následně využit pro syntézu aminokyselin, proteinů, nukleových kyselin a dalších sloučenin obsahujících dusík. Bakterie uzlíků a luštěniny se navzájem potřebují. Společná oboustranně výhodná existence odlišné typy organismům se říká symbióza.
Kromě fotoautotrofů jsou některé bakterie (vodíkové, nitrifikační, sirné bakterie aj.) schopny syntetizovat organické látky z látek anorganických. Tuto syntézu provádějí díky energii uvolněné při oxidaci anorganických látek. Říká se jim chemoautotrofy. Proces chemosyntézy objevil v roce 1887 ruský mikrobiolog S. N. Vinogradsky.
Všechny živé bytosti naší planety, které nejsou schopny syntetizovat organické látky z anorganických sloučenin, se nazývají heterotrofy. Všichni živočichové i lidé žijí z energie Slunce uložené rostlinami, přeměněné na energii chemických vazeb nově syntetizovaných organických sloučenin.
Je třeba poznamenat, že fotosyntetické i chemosyntetické organismy jsou také schopny získávat energii díky oxidaci organických látek. Heterotrofní však tyto látky přijímají zvenčí hotové, autotrofní je syntetizují z anorganických sloučenin.
Fotosyntetické buňky absorbují oxid uhličitý z atmosféry a uvolňují do ní kyslík. Před příchodem fotosyntetických buněk na naši planetu byla zemská atmosféra bez kyslíku. S nástupem fotosyntetických organismů vedlo postupné zaplňování atmosféry kyslíkem ke vzniku buněk s novým typem energetického aparátu. Jednalo se o buňky, které vyráběly energii oxidací hotových organických sloučenin, hlavně sacharidů a tuků, za účasti vzdušného kyslíku jako oxidačního činidla. Při oxidaci organických sloučenin se uvolňuje energie.
V důsledku nasycení atmosféry kyslíkem vznikly aerobní buňky, které jsou schopny využívat kyslík k energii.
§ 11. Fotosyntéza. Přeměna světelné energie na energii chemické vazby
První buňky schopné využívat energii slunečního světla se na Zemi objevily asi před 4 miliardami let v éře Archeanů. Jednalo se o sinice (z řeckého „cyanos“ – modrá). Jejich zkamenělé pozůstatky byly nalezeny v břidlicových vrstvách pocházejících z tohoto období historie Země. Dalších 1,5 miliardy let trvalo nasycení zemské atmosféry kyslíkem a vznik aerobních buněk.
Je zřejmé, že role rostlin a dalších fotosyntetických organismů ve vývoji a udržování života na naší planetě je mimořádně velká: přeměňují energii slunečního záření na energii chemických vazeb organických sloučenin, kterou pak využívají všechny ostatní živé organismy. bytosti; nasycují atmosféru Země kyslíkem, který slouží k oxidaci organických látek a tím získávání chemické energie v nich uložené aerobními buňkami; Konečně některé rostlinné druhy v symbióze s bakteriemi fixujícími dusík zavádějí plynný dusík z atmosféry do molekul amoniaku, jeho solí a organických sloučenin obsahujících dusík.
Roli zelených rostlin v planetárním životě nelze přeceňovat. Zachování a rozšiřování zeleného pokryvu Země má zásadní význam pro všechny živé bytosti obývající naši planetu.
Ukládání světelné energie do biologických „akumulátorů“. Proud slunečního světla nese světelné vlny různých délek. Rostliny pomocí světelných „antén“ (jedná se především o molekuly chlorofylu) pohlcují světelné vlny červené a modré části spektra. Chlorofyl prochází světelné vlny zelené části spektra bez zpoždění, a proto mají rostliny zelenou barvu.
Pomocí světelné energie se elektron ve složení molekuly chlorofylu přenese na vyšší energetickou hladinu. Dále tento vysokoenergetický elektron, jako by na schodech, skáče podél řetězce nosičů elektronů a ztrácí energii. V tomto případě se energie elektronů vynakládá na „nabíjení“ jakési biologické „akumulátory“. Aniž bychom se pouštěli do chemických vlastností jejich struktury, řekněme, že jednou z nich je kyselina adenosintrifosforečná, která se také nazývá adenosintrifosfát (zkráceně ATP). Jak již bylo uvedeno v § 6, ATP obsahuje tři navzájem spojené zbytky kyseliny fosforečné, které jsou navázány na adenosin. Schematicky lze ATP popsat vzorcem: adenosin-P-P~P, kde P je zbytek kyseliny fosforečné. V chemické vazbě mezi druhým a třetím koncovým fosfátem je uložena energie, kterou elektron odevzdá (taková zvláštní chemická vazba je znázorněna vlnovkou). To je způsobeno tím, že když elektron předá svou energii adenosindifosfátu (adenosin-FF, ADP), přidá se další fosfát: ADP + P + E → ATP, kde E je energie elektronu, která je uložena v ATP. Při štěpení ATP enzymem adenosintrifosfatázou (ATPáza) se odštěpí koncový fosfát a uvolní se energie:
V rostlinné buňce se energie ATP využívá k transportu vody a solí, k buněčnému dělení, růstu a pohybu (vzpomeňte si, jak se otáčí hlava slunečnice za Sluncem).
Energie ATP je nezbytná pro syntézu glukózy, škrobu, celulózy a dalších organických sloučenin v rostlinách. Pro syntézu organických látek v rostlinách je však potřeba ještě jeden biologický „akumulátor“, který uchovává energii světla. Tato baterie má nevyslovitelný dlouhý název: nikotinamid adenindinukleotid fosfát (zkráceně NADP, vyslovováno „nad-ef“). Tato sloučenina existuje v redukované, vysokoenergetické formě: NADP-N (vyslovuje se „nad-ef-ash“).
Oxidovaná forma této sloučeniny je NADP+ (vyslovuje se „nad-ef-plus“). Ztrátou jednoho atomu vodíku a jednoho elektronu se NADP-N mění na NADP + a redukuje oxid uhličitý (za účasti molekul vody) na glukózu C 6 H 12 0 6; chybějící protony (H +) jsou odebírány z vodního prostředí. Ve zjednodušené formě lze tento proces zapsat jako chemickou rovnici:
Při smíchání oxidu uhličitého a vody však glukóza nevzniká. K tomu je zapotřebí nejen obnovující síla NADPH, ale také energie ATP a sloučeniny, která váže CO 2, který se využívá v mezistupních syntézy glukózy, a také řada enzymů – biologických katalyzátorů tohoto procesu.
Fotolýza vody. Jak vzniká kyslík během fotosyntézy? Energie světla se totiž spotřebuje i na štěpení molekuly vody – fotolýzu. V tomto případě se tvoří protony (H +), elektrony (O a volný kyslík:
Elektrony vzniklé při fotolýze kompenzují jejich ztrátu chlorofylem (jak se říká, vyplňují „díru“, která v chlorofylu vznikla). Část elektronů za účasti protonů redukuje NADP + na NADP-H. Vedlejším produktem této reakce je kyslík (obr. 19). Jak je vidět ze souhrnné rovnice pro syntézu glukózy, uvolňuje se při tomto procesu kyslík.
Když rostliny využívají energii slunečního světla, nepotřebují kyslík. Při nedostatku slunečního světla se však rostliny stávají aerobními. V noční tmě spotřebovávají kyslík a oxidují glukózu, fruktózu, škrob a další sloučeniny uložené během dne jako zvířata.
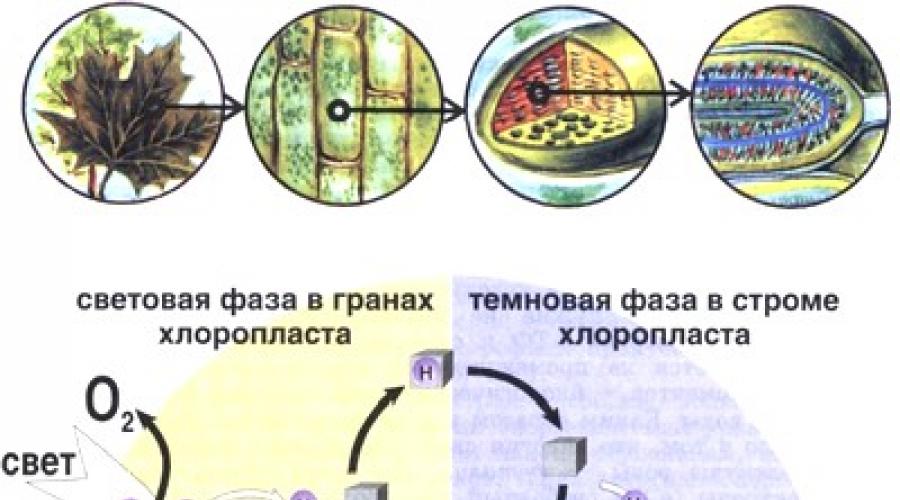
Světlé a tmavé fáze fotosyntézy. V procesu fotosyntézy se rozlišují světlé a tmavé fáze. Při osvětlení rostlin se energie světla přemění na energii chemických vazeb ATP a NADP-H. Energie těchto sloučenin se snadno uvolňuje a využívá uvnitř rostlinné buňky pro různé účely, především pro syntézu glukózy a dalších organických sloučenin. Proto se tato počáteční fáze fotosyntézy nazývá světelná fáze. Bez osvětlení slunečním nebo umělým světlem, v jehož spektru jsou červené a modré paprsky, nedochází v rostlinné buňce k syntéze ATP a NADP-H. Když se však molekuly ATP a NADP-H již nahromadily v rostlinné buňce, může k syntéze glukózy dojít i ve tmě, bez účasti světla. Pro tyto biochemické reakce není potřeba osvětlení, protože jsou již vybaveny světelnou energií uloženou v biologických „akumulátorech“. Tato fáze fotosyntézy se nazývá teplotní fáze.

Rýže. 19. Schéma fotosyntézy
Všechny reakce fotosyntézy probíhají v chloroplastech - ztluštělé oválné nebo kulaté útvary umístěné v cytoplazmě rostlinné buňky (o chloroplastech již bylo krátce pojednáno v § 9). Každá buňka obsahuje 40-50 chloroplastů. Chloroplasty jsou zvenku ohraničeny dvojitou membránou a uvnitř jsou tenké ploché váčky - tylakoidy, rovněž ohraničené membránami. Thylakoidy obsahují chlorofyl, nosiče elektronů a všechny enzymy podílející se na světelné fázi fotosyntézy, dále ADP, ATP, NADP + a NADP-H. Desítky thylakoidů jsou pevně zabaleny do stohů zvaných grana. v vnitřní prostor mezi granou - ve stromatu chloroplastů - jsou enzymy podílející se na redukci CO 2 na glukózu vlivem energie produktů světelné fáze fotosyntézy - ATP a NADP-H. V důsledku toho dochází ve stromatu k reakcím temné fáze fotosyntézy, úzce souvisejících se světlou fází, která se rozvíjí v thylakoidech. Světlá a temná fáze fotosyntézy jsou schematicky znázorněny na obrázku 19.
Chloroplasty mají svůj vlastní genetický aparát – molekuly DNA a autonomně se množí uvnitř buněk. Předpokládá se, že před více než 1,5 miliardami let to byly volné mikroorganismy, které se staly symbionty rostlinných buněk.
- Vysvětlete, proč říkáme, že energii pro život na Zemi zpočátku dodává Slunce.
- Vysvětlete, proč se při fotosyntéze využívá oxid uhličitý a voda a co je zdrojem vedlejšího produktu fotosyntézy, kyslíku.
- Jak souvisí problémy fotosyntézy a zásobování obyvatel Země potravou?
- Proč fotosyntéza přeměňuje energii slunečního záření dopadajícího na list na energii uloženou v organických sloučeninách s účinností jen asi 1 %? Jaký je osud zbytku energie?
- Vyplňte tabulku.
Historie studia fotosyntézy sahá do srpna 1771, kdy anglický teolog, filozof a amatérský přírodovědec Joseph Priestley (1733-1804) zjistil, že rostliny dokážou „opravit“ vlastnosti vzduchu, který mění své složení v důsledku spalování resp. život zvířat. Priestley ukázal, že v přítomnosti rostlin se „zkažený“ vzduch opět stává vhodným ke spalování a podpoře života zvířat.
V průběhu dalších studií Ingenhause, Senebiera, Saussura, Bussenga a dalších vědců bylo zjištěno, že rostliny, když jsou osvětleny, uvolňují kyslík a absorbují oxid uhličitý ze vzduchu. Rostliny syntetizují organické látky z oxidu uhličitého a vody. Tento proces se nazýval fotosyntéza.
Robert Mayer, který objevil zákon zachování energie, navrhl v roce 1845, že rostliny přeměňují energii slunečního světla na energii chemických sloučenin vznikajících během fotosyntézy. Podle něj jsou "sluneční paprsky šířící se vesmírem "zachycovány" a ukládány pro další použití podle potřeby." Následně ruský vědec K.A. Timiryazev přesvědčivě dokázal, že nejdůležitější roli ve využívání sluneční energie rostlinami hrají molekuly chlorofylu přítomné v zelených listech.
Sacharidy (cukry) vznikající při fotosyntéze se využívají jako zdroj energie a stavební materiál pro syntézu různých organických sloučenin v rostlinách a zvířatech. U vyšších rostlin probíhají procesy fotosyntézy v chloroplastech – specializovaných organelách rostlinné buňky přeměňujících energii.
Schematické znázornění chloroplastu je znázorněno na Obr. jeden.
Pod dvojitým obalem chloroplastu, který se skládá z vnější a vnitřní membrány, jsou rozšířené membránové struktury, které tvoří uzavřené váčky zvané tylakoidy. Tylakoidní membrány se skládají ze dvou vrstev lipidových molekul, které zahrnují makromolekulární fotosyntetické proteinové komplexy. V chloroplastech vyšších rostlin jsou thylakoidy seskupeny do grana, což jsou stohy diskovitých, zploštělých a těsně k sobě přitlačených thylakoidů. Z nich vyčnívající intergranální thylakoidy jsou pokračováním jednotlivých thylakoidů grana. Prostor mezi chloroplastovou membránou a tylakoidy se nazývá stroma. Stroma obsahuje RNA, DNA, molekuly chloroplastů, ribozomy, škrobová zrna a četné enzymy, včetně těch, které zajišťují příjem CO2 rostlinami.
Publikace vznikla s podporou Sushi E'xpress. Společnost Sushi E'xpress poskytuje služby rozvozu sushi v Novosibirsku. Objednáním sushi ze Sushi E’xpress rychle získáte chutný a zdravý pokrm vyrobený profesionálními kuchaři z těch nejčerstvějších produktů té nejvyšší kvality. Návštěvou webu Sushi E’xpress se můžete seznámit s cenami a složením nabízených rohlíků, které vám pomohou při rozhodování při výběru pokrmu. Chcete-li zadat objednávku na doručení sushi, volejte 239-55-87
Světlé a tmavé fáze fotosyntézy
Podle moderní nápady, fotosyntéza je řada fotofyzikálních a biochemických procesů, v jejichž důsledku rostliny syntetizují sacharidy (cukry) pomocí energie slunečního světla. Četné fáze fotosyntézy se obvykle dělí na dvě velké skupiny procesů – světlou a tmavou fázi.
Světelná stádia fotosyntézy je zvykem nazývat souborem procesů, v jejichž důsledku se vlivem energie světla syntetizují molekuly adenosintrifosfátu (ATP) a vzniká redukovaný nikotinamid adenindinukleotidfosfát (NADPH). sloučenina s vysokým redukčním potenciálem. Molekuly ATP fungují v buňce jako univerzální zdroj energie. Je známo, že energie vysokoenergetických (tj. energeticky bohatých) fosfátových vazeb molekuly ATP se využívá ve většině biochemických procesů náročných na energii.
Světelné procesy fotosyntézy probíhají v thylakoidech, jejichž membrány obsahují hlavní složky fotosyntetického aparátu rostlin - komplexy světlosběrného pigmentu a proteinu a elektronového transportu a také komplex ATP-syntázy, který katalyzuje tvorbu ATP. z adenosindifosfátu (ADP) a anorganického fosfátu (F i) (ADP + F i → ATP + H 2 O). Energie světla absorbovaná rostlinami se tedy v důsledku světelných fází fotosyntézy ukládá ve formě makroergických chemických vazeb molekul ATP a silného redukčního činidla NADP H, které se používají k syntéze sacharidů v tzv. temné fáze fotosyntézy.
Temná stádia fotosyntézy se obvykle označují jako soubor biochemických reakcí, jejichž výsledkem je asimilace atmosférického oxidu uhličitého (CO 2) rostlinami a tvorba sacharidů. Cyklus temných biochemických přeměn vedoucích k syntéze organických sloučenin z CO2 a vody se nazývá Calvin-Bensonův cyklus podle jmen autorů, kteří rozhodujícím způsobem přispěli ke studiu těchto procesů. Na rozdíl od transportu elektronů a komplexů ATP syntázy, které se nacházejí v thylakoidní membráně, jsou enzymy, které katalyzují „temné“ reakce fotosyntézy, rozpuštěny ve stromatu. Při zničení chloroplastové membrány jsou tyto enzymy vymyty ze stromatu, v důsledku čehož chloroplasty ztrácejí schopnost absorbovat oxid uhličitý.
V důsledku přeměn řady organických sloučenin v Calvin-Bensonově cyklu vzniká ze tří molekul CO 2 a vody v chloroplastech molekula glyceraldehyd-3-fosfátu, která má chemický vzorec CHO–CHOH–CH 2 O– PO 3 2-. Na jednu molekulu CO 2 obsaženou v glyceraldehyd-3-fosfátu se přitom spotřebují tři molekuly ATP a dvě molekuly NADP H.
Pro syntézu organických sloučenin v Calvin-Bensonově cyklu se využívá energie uvolněná při hydrolýze makroergických fosfátových vazeb molekul ATP (reakce ATP + H 2 O → ADP + F i) a silný redukční potenciál molekul NADP H. Hlavní část molekul vytvořených v chloroplastu Glyceraldehyd-3-fosfát vstupuje do cytosolu rostlinné buňky, kde se přeměňuje na fruktóza-6-fosfát a glukóza-6-fosfát, které v průběhu dalších přeměn tvoří sacharofosfát, prekurzor sacharózy. Z molekul glyceraldehyd-3-fosfátu zbývajících v chloroplastu se syntetizuje škrob.
Přeměna energie ve fotosyntetických reakčních centrech
Fotosyntetické komplexy rostlin, řas a fotosyntetických bakterií, které přeměňují energii, byly dobře studovány. Instalováno chemické složení a prostorové struktury proteinových komplexů přeměňujících energii, byla objasněna sekvence procesů přeměny energie. Navzdory rozdílům ve složení a molekulární struktuře fotosyntetického aparátu existují společné vzorce v procesech přeměny energie ve fotoreakčních centrech všech fotosyntetických organismů. Ve fotosyntetických systémech rostlinného i bakteriálního původu je jediným strukturálním a funkčním článkem fotosyntetického aparátu. fotosystém, jehož součástí je světlosběrná anténa, centrum fotochemické reakce a s ním spojené molekuly – nosiče elektronů.
Nejprve zvažte obecné zásady přeměna sluneční energie, charakteristická pro všechny fotosyntetické systémy, a dále se podrobněji zastavíme na příkladu fungování fotoreakčních center a elektronového transportního řetězce chloroplastů ve vyšších rostlinách.
Anténa pro sběr světla (absorpce světla, migrace energie do reakčního centra)
Úplně prvním elementárním aktem fotosyntézy je absorpce světla molekulami chlorofylu nebo pomocnými pigmenty, které jsou součástí speciálního pigment-proteinového komplexu zvaného světlosběrná anténa. Anténa pro sběr světla je makromolekulární komplex navržený k efektivnímu zachycování světla. V chloroplastech obsahuje anténní komplex velké množství (až několik stovek) molekul chlorofylu a určité množství pomocných pigmentů (karotenoidy) silně spojených s proteinem.
Při jasném slunečním světle absorbuje jedna molekula chlorofylu světelná kvanta poměrně zřídka, v průměru ne více než 10krát za sekundu. Protože však na jedno fotoreakční centrum připadá velký počet molekul chlorofylu (200–400), dochází i při relativně nízké intenzitě světla dopadajícího na list za podmínek stínování k aktivaci reakčního centra poměrně často. Soubor pigmentů absorbujících světlo ve skutečnosti hraje roli antény, která díky své poměrně velké velikosti účinně zachycuje sluneční světlo a směruje jeho energii do reakčního centra. Stínomilné rostliny obvykle mají větší velikost anténa pro sklizeň světla ve srovnání s rostlinami rostoucími ve vysokých světelných podmínkách.
Molekuly chlorofylu jsou hlavními světlosběrnými pigmenty v rostlinách. A a chlorofyl b absorbující viditelné světlo o vlnové délce λ ≤ 700–730 nm. Izolované molekuly chlorofylu absorbují světlo pouze ve dvou relativně úzkých pásmech slunečního spektra: na vlnových délkách 660–680 nm (červené světlo) a 430–450 nm (modrofialové světlo), což samozřejmě omezuje efektivitu využití celé spektrum slunečního světla dopadajícího na zelený list.
Spektrální složení světla absorbovaného světlosběrnou anténou je však ve skutečnosti mnohem širší. To se vysvětluje tím, že absorpční spektrum agregovaných forem chlorofylu, které jsou součástí světlosběrné antény, se posouvá směrem k delším vlnovým délkám. Spolu s chlorofylem jsou součástí světlosběrné antény pomocné pigmenty, které zvyšují její účinnost díky tomu, že absorbují světlo v těch spektrálních oblastech, ve kterých molekuly chlorofylu, hlavního pigmentu světlosběrné antény, světlo absorbují poměrně slabě.
U rostlin jsou pomocnými pigmenty karotenoidy, které absorbují světlo v rozsahu vlnových délek λ ≈ 450–480 nm; v buňkách fotosyntetických řas jsou to červené a modré pigmenty: fykoerytriny u červených řas (λ ≈ 495–565 nm) a fykocyaniny u modrozelených řas (λ ≈ 550–615 nm).
Absorpce světelného kvanta molekulou chlorofylu (Chl) nebo pomocným pigmentem vede k jeho excitaci (elektron přejde na vyšší energetickou hladinu):
Chl + hν → Chl*.
Energie excitované molekuly chlorofylu Chl* se přenáší na molekuly sousedních pigmentů, které ji zase mohou přenášet na další molekuly světlosběrné antény:
Chl* + Chl → Chl + Chl*.
 Excitační energie tak může migrovat skrz pigmentovou matrici, dokud excitace nakonec nedosáhne fotoreakčního centra P (schematické znázornění tohoto procesu je na obr. 2):
Excitační energie tak může migrovat skrz pigmentovou matrici, dokud excitace nakonec nedosáhne fotoreakčního centra P (schematické znázornění tohoto procesu je na obr. 2):
Chl* + P → Chl + P*.
Všimněte si, že doba existence molekul chlorofylu a dalších pigmentů v excitovaném stavu je velmi krátká, τ ≈ 10–10–10–9 s. Je tedy jistá pravděpodobnost, že na cestě do reakčního centra P se energie takto krátkodobě excitovaných stavů pigmentů může zbytečně ztratit - rozptýlit do tepla nebo uvolnit ve formě světelného kvanta (fenomén fluorescence). Ve skutečnosti je však účinnost migrace energie do fotosyntetického reakčního centra velmi vysoká. V případě, že je reakční centrum v aktivním stavu, je pravděpodobnost ztráty energie zpravidla maximálně 10–15 %. Tak vysoká účinnost využití energie slunečního světla je způsobena tím, že světlosběrná anténa je vysoce uspořádaná struktura, která zajišťuje velmi dobrou vzájemnou interakci pigmentů. Díky tomu je dosaženo vysoké rychlosti přenosu excitační energie z molekul absorbujících světlo do fotoreakčního centra. Průměrná doba „skoku“ excitační energie z jednoho pigmentu na druhý je zpravidla τ ≈ 10–12–10–11 s. Celková doba migrace excitace do reakčního centra obvykle nepřesahuje 10–10–10–9 s.
Centrum fotochemické reakce (přenos elektronů, stabilizace separovaných nábojů)
Moderním představám o struktuře reakčního centra a mechanismech primárních fází fotosyntézy předcházely práce A.A. Krasnovského, který objevil, že v přítomnosti donorů a akceptorů elektronů mohou být molekuly chlorofylu excitované světlem reverzibilně redukovány (přijmout elektron) a oxidovat (darovat elektron). Následně v rostlinách, řasách a fotosyntetických bakteriích Kok, Witt a Duizens objevili speciální pigmenty chlorofylové povahy zvané reakční centra, která se působením světla oxidují a jsou vlastně primárními donory elektronů při fotosyntéze.
Fotochemické reakční centrum P je speciální pár (dimer) molekul chlorofylu, které působí jako past na excitační energii putující pigmentovou matricí světlosběrné antény (obr. 2). Stejně jako kapalina stéká ze stěn široké nálevky k jejímu úzkému hrdlu, energie světla absorbovaného všemi pigmenty světlosběrné antény směřuje do reakčního centra. Excitace reakčního centra iniciuje řetězec dalších přeměn světelné energie během fotosyntézy.
Sled procesů probíhajících po excitaci reakčního centra P a diagram odpovídajících změn energie fotosystému jsou schematicky znázorněny na Obr. 3.
Spolu s dimerem chlorofylu P obsahuje fotosyntetický komplex molekuly primárních a sekundárních akceptorů elektronů, které budeme konvenčně označovat symboly A a B, a také primární donor elektronů, molekulu D. k sousednímu primárnímu akceptoru elektronů A :
D(P*A)B → D(P + A –)B.
V důsledku velmi rychlého (t ≈ 10–12 s) přenosu elektronů z P* do A je tedy realizován druhý zásadně důležitý krok přeměny sluneční energie při fotosyntéze – separace náboje v reakčním centru. V tomto případě vzniká silné redukční činidlo A - (donor elektronu) a silné oxidační činidlo P + (akceptor elektronu).
Molekuly P + a A - jsou umístěny v membráně asymetricky: v chloroplastech je reakční centrum P + blíže k povrchu membrány směřující dovnitř thylakoidu a akceptor A - je umístěn blíže k vnější straně. V důsledku fotoindukované separace náboje proto na membráně vzniká rozdíl v elektrických potenciálech. Světlem indukovaná separace nábojů v reakčním centru je podobná generování rozdílu elektrického potenciálu v konvenčním fotočlánku. Je však třeba zdůraznit, že na rozdíl od všech známých a v technologii široce používaných energetických fotokonvertorů je účinnost fotosyntetických reakčních center velmi vysoká. Účinnost separace náboje v aktivních fotosyntetických reakčních centrech zpravidla přesahuje 90–95 % (u nejlepších vzorků fotočlánků účinnost nepřesahuje 30 %).
Jaké mechanismy zajišťují tak vysokou účinnost přeměny energie v reakčních centrech? Proč se elektron přenesený na akceptor A nevrátí zpět do kladně nabitého oxidovaného centra P +? Stabilizace separovaných nábojů je zajištěna především sekundárními procesy transportu elektronů po přenosu elektronu z P* do A. Z redukovaného primárního akceptoru A se elektron velmi rychle (za 10–10–10–9 s ) jde na akceptor sekundárních elektronů B:
D(P + A –)B → D(P + A)B – .
V tomto případě dochází nejen k odstranění elektronu z kladně nabitého reakčního centra P +, ale znatelně klesá i energie celého systému (obr. 3). To znamená, že k přenosu elektronu v opačném směru (přechod B – → A) bude muset překonat dostatečně vysokou energetickou bariéru ΔE ≈ 0,3–0,4 eV, kde ΔE je rozdíl energetických hladin pro dva stavy systém, ve kterém je elektron na nosiči A nebo B. Z tohoto důvodu by návrat elektronu zpět, z redukované molekuly B do oxidované molekuly A, trval mnohem déle než u přímého přechodu A - → B. Jinými slovy, v dopředném směru se elektron přenáší mnohem rychleji než naopak. Po přenosu elektronu na sekundární akceptor B se tedy výrazně snižuje pravděpodobnost jeho návratu zpět a rekombinace s kladně nabitou „dírou“ P +.
Druhým faktorem přispívajícím ke stabilizaci separovaných nábojů je rychlá neutralizace oxidovaného fotoreakčního centra P+ vlivem elektronu přicházejícího do P+ z elektronového donoru D:
D(P + A)B – → D + (PA)B – .
Po přijetí elektronu z donorové molekuly D a návratu do původního redukovaného stavu P, reakční centrum již nebude schopno přijmout elektron z redukovaných akceptorů, ale nyní je připraveno znovu spustit - darovat elektron vedle něj se nachází oxidovaný primární akceptor A. Jedná se o sled dějů, které probíhají ve fotoreakčních centrech všech fotosyntetických systémů.
Elektronový transportní řetězec chloroplastu
V chloroplastech vyšších rostlin jsou dva fotosystémy: fotosystém 1 (PS1) a fotosystém 2 (PS2), které se liší složením bílkovin, pigmentů a optické vlastnosti. Světlosběrná anténa PS1 absorbuje světlo o vlnové délce λ ≤ 700–730 nm a PS2 absorbuje světlo o vlnové délce λ ≤ 680–700 nm. Světlem indukovaná oxidace reakčních center PS1 a PS2 je doprovázena jejich odbarvením, které je charakterizováno změnami jejich absorpčních spekter při λ ≈ 700 a 680 nm. V souladu s jejich optickými charakteristikami byla reakční centra PS1 a PS2 pojmenována P 700 a P 680.
Oba fotosystémy jsou propojeny řetězem elektronových nosičů (obr. 4). PS2 je zdrojem elektronů pro PS1. Světlem iniciovaná separace náboje ve fotoreakčních centrech P 700 a P 680 zajišťuje přenos elektronů z vody rozložené v PS2 do konečného akceptoru elektronů, molekuly NADP+. Elektronový transportní řetězec (ETC) spojující dva fotosystémy zahrnuje molekuly plastochinonu, samostatný elektronový transportní proteinový komplex (tzv. b/f komplex) a ve vodě rozpustný protein plastocyanin (Pc) jako nosiče elektronů. Schéma znázorňující vzájemné uspořádání elektronových transportních komplexů v thylakoidní membráně a dráhu přenosu elektronů z vody do NADP + je znázorněno na Obr. čtyři.
V PS2 je elektron přenesen z excitovaného centra P * 680 nejprve do primárního akceptorového feofetinu (Phe) a poté do plastochinonové molekuly Q A, pevně vázané na jeden z proteinů PS2,
Y(P* 680 Phe)Q A Q B → Y(P + 680 Phe –)Q A Q B → Y(P + 680 Phe)Q A – Q B .
Poté se elektron přenese na druhou molekulu plastochinonu Q B a P 680 přijme elektron od primárního donoru elektronu Y:
Y(P + 680 Phe)Q A – Q B → Y + (P 680 Phe)Q A Q B – .
 Molekula plastochinonu, jejíž chemický vzorec a umístění v lipidové dvouvrstvé membráně je znázorněno na Obr. 5 je schopen přijmout dva elektrony. Poté, co se reakční centrum PS2 spustí dvakrát, molekula plastochinonu Q B obdrží dva elektrony:
Molekula plastochinonu, jejíž chemický vzorec a umístění v lipidové dvouvrstvé membráně je znázorněno na Obr. 5 je schopen přijmout dva elektrony. Poté, co se reakční centrum PS2 spustí dvakrát, molekula plastochinonu Q B obdrží dva elektrony:
Q B + 2е – → Q B 2– .
Záporně nabitá molekula Q B 2– má vysokou afinitu k vodíkovým iontům, které zachycuje ze stromálního prostoru. Po protonaci redukovaného plastochinonu Q B 2– (Q B 2– + 2H + → QH 2) vzniká elektricky neutrální forma této molekuly QH 2, která se nazývá plastochinol (obr. 5). Plastochinol hraje roli mobilního nosiče dvou elektronů a dvou protonů: po opuštění PS2 se molekula QH2 může snadno pohybovat uvnitř thylakoidní membrány, čímž poskytuje spojení mezi PS2 a dalšími komplexy transportu elektronů.
Oxidované reakční centrum PS2 P 680 má výjimečně vysokou elektronovou afinitu; je velmi silné oxidační činidlo. Díky tomu se voda, chemicky stabilní sloučenina, v PS2 rozkládá. Water splitting complex (WRC) obsažený v PS2 obsahuje ve svém aktivním středu skupinu manganových iontů (Mn 2+), které slouží jako elektronové donory pro P 680 . Darováním elektronů do oxidovaného reakčního centra se ionty manganu stávají „akumulátory“ kladných nábojů, které se přímo účastní oxidační reakce vody. V důsledku postupné čtyřnásobné aktivace reakčního centra P 680 se v aktivním centru WRC obsahujícím Mn hromadí čtyři silné oxidační ekvivalenty (neboli čtyři „otvory“) ve formě oxidovaných iontů manganu (Mn 4+). , které interakcí se dvěma molekulami vody katalyzují rozkladnou reakční vodu:
2Mn4+ + 2H20 -> 2Mn2+ + 4H++02.
Po postupném přenosu čtyř elektronů z WRC do P 680 tedy dochází k synchronnímu rozkladu dvou molekul vody najednou, doprovázeném uvolněním jedné molekuly kyslíku a čtyř vodíkových iontů, které se dostanou do intratylakoidního prostoru chloroplastu.
Molekula plastochinolu QH2 vzniklá během fungování PS2 difunduje do lipidové dvojvrstvy thylakoidní membrány ke komplexu b/f (obr. 4 a 5). Při srážce s komplexem b/f se na něj naváže molekula QH 2 a následně na něj přenese dva elektrony. V tomto případě se na každou molekulu plastochinolu zoxidovanou komplexem b/f uvolní dva vodíkové ionty uvnitř thylakoidu. Komplex b/f zase slouží jako donor elektronů pro plastocyanin (Pc), relativně malý ve vodě rozpustný protein, jehož aktivní centrum obsahuje ionty mědi (redukční a oxidační reakce plastocyaninu jsou doprovázeny změnami v mocenství mědi iont Cu 2+ + e – ↔Cu+). Plastocyanin působí jako spojovací článek mezi b/f komplexem a PS1. Molekula plastocyaninu se v thylakoidu rychle pohybuje a zajišťuje přenos elektronů z komplexu b/f do PS1. Z redukovaného plastocyaninu jde elektron přímo do oxidovaných reakčních center PS1 – P 700 + (viz obr. 4). V důsledku společného působení PS1 a PS2 se tedy dva elektrony z molekuly vody rozložené v PS2 nakonec přenesou přes elektronový transportní řetězec do molekuly NADP +, čímž se vytvoří silné redukční činidlo NADP H.
Proč chloroplasty potřebují dva fotosystémy? Je známo, že fotosyntetické bakterie, které využívají různé organické a anorganické sloučeniny (například H 2 S) jako donor elektronů k redukci oxidovaných reakčních center, úspěšně fungují s jedním fotosystémem. Vznik dvou fotosystémů je pravděpodobně způsoben tím, že energie jednoho kvanta viditelného světla nestačí k zajištění rozkladu vody a efektivního průchodu elektronu po celé délce řetězce nosných molekul z vody do NADP. + . Zhruba před 3 miliardami let se na Zemi objevily modrozelené řasy nebo sinice, které získaly schopnost využívat vodu jako zdroj elektronů k redukci oxidu uhličitého. Nyní se předpokládá, že PS1 pochází ze zelených bakterií a PS2 z purpurových bakterií. Poté, co byl PS2 během evolučního procesu „zahrnut“ do jediného elektronového transportního řetězce spolu s PS1, bylo možné vyřešit energetický problém - překonat poměrně velký rozdíl v redox potenciálu párů kyslík / voda a NADP + / NADP H. Jedním z nich se stal vznik fotosyntetických organismů, schopných oxidovat vodu milníky vývoj divoké zvěře na Zemi. Za prvé, řasy a zelené rostliny, které se „naučily“ oxidovat vodu, ovládly nevyčerpatelný zdroj elektronů pro redukci NADP+. Za druhé, rozkladem vody naplnily zemskou atmosféru molekulárním kyslíkem a vytvořily tak podmínky pro rychlý evoluční vývoj organismů, jejichž energie je spojena s aerobním dýcháním.
Spojení procesů transportu elektronů s přenosem protonů a syntézou ATP v chloroplastech
Přenos elektronu po CET je zpravidla doprovázen úbytkem energie. Tento proces lze přirovnat ke spontánnímu pohybu tělesa po nakloněné rovině. Pokles energetické hladiny elektronu při jeho pohybu po CET vůbec neznamená, že přenos elektronu je vždy energeticky zbytečný proces. Za normálních podmínek fungování chloroplastů většina energie uvolněné při transportu elektronů nepřijde nazmar, ale využije se k provozu speciálního komplexu přeměňujícího energii zvaného ATP syntáza. Tento komplex katalyzuje energeticky nepříznivý proces tvorby ATP z ADP a anorganického fosfátu F i (reakce ADP + F i → ATP + H 2 O). V tomto ohledu je zvykem říkat, že procesy přenosu elektronů poskytující energii jsou spojeny s procesy syntézy ATP přijímajícími energii.
Procesy transportu protonů hrají nejdůležitější roli při zajišťování energetické konjugace v thylakoidních membránách, stejně jako ve všech ostatních organelách přeměňujících energii (mitochondrie, chromatofory fotosyntetických bakterií). Syntéza ATP úzce souvisí s přenosem tří protonů přes ATP syntázu z thylakoidů (3H in +) do stromatu (3H out +):
ADP + F i + 3H in + → ATP + H20 + 3H out +.
Tento proces je možný, protože v důsledku asymetrického uspořádání nosičů v membráně vede fungování chloroplastu ETC k akumulaci nadměrného množství protonů uvnitř thylakoidu: vodíkové ionty jsou absorbovány zvenčí ve fázích NADP + redukce a tvorbu plastochinolu a jsou uvolňovány uvnitř thylakoidů ve stádiích rozkladu vody a oxidace plastochinolu (obr. 4). Osvětlení chloroplastů vede k výraznému (100–1000násobnému) zvýšení koncentrace vodíkových iontů uvnitř thylakoidů.
Uvažovali jsme tedy o řetězci dějů, během kterého se energie slunečního světla ukládá ve formě energie vysokoenergetických chemických sloučenin - ATP a NADP H. Tyto produkty světelné fáze fotosyntézy se používají v temných fázích k tvorbě organické sloučeniny (sacharidy) z oxidu uhličitého a vody. Hlavní kroky přeměny energie, vedoucí ke vzniku ATP a NADP H, zahrnují následující procesy: 1) absorpce světelné energie pigmenty světlosběrné antény; 2) přenos excitační energie do centra fotoreakce; 3) oxidace centra fotoreakce a stabilizace separovaných nábojů; 4) přenos elektronů podél řetězce přenosu elektronů, tvorba NADP H; 5) transmembránový přenos vodíkových iontů; 6) Syntéza ATP.
1. Alberts B., Bray D., Lewis J., Roberts K., Watson J. Molekulární biologie buňky. T. 1. - M.: Mir, 1994. 2. vyd.
2. Kukushkin A.K., Tichonov A.N. Přednášky o biofyzice rostlinné fotosyntézy. – M.: Nakladatelství Moskevské státní univerzity, 1988.
3. Nichols D.D. Bioenergetika. Úvod do chemiosmotické teorie. – M.: Mir, 1985.
4. Skulachev V.P. Energie biologických membrán. – M.: Nauka, 1989.
Jak se energie slunečního světla ve světlé a tmavé fázi fotosyntézy přeměňuje na energii chemických vazeb glukózy? Vysvětlete odpověď.
Odpovědět
Ve světelné fázi fotosyntézy se energie slunečního světla přeměňuje na energii excitovaných elektronů a následně se energie excitovaných elektronů přeměňuje na energii ATP a NADP-H2. V temné fázi fotosyntézy se energie ATP a NADP-H2 přeměňuje na energii chemických vazeb glukózy.
Co se děje během světelné fáze fotosyntézy?
Odpovědět
Elektrony chlorofylu, excitované energií světla, jdou podél elektronových transportních řetězců, jejich energie je uložena v ATP a NADP-H2. Dochází k fotolýze vody, uvolňuje se kyslík.
Jaké jsou hlavní procesy, které probíhají během temné fáze fotosyntézy?
Odpovědět
Z oxidu uhličitého získaného z atmosféry a vodíku získaného ve světelné fázi vzniká glukóza díky energii ATP získané ve světelné fázi.
Jaká je funkce chlorofylu v rostlinné buňce?
Odpovědět
Chlorofyl se podílí na procesu fotosyntézy: ve světelné fázi chlorofyl absorbuje světlo, elektron chlorofylu přijímá světelnou energii, odlomí se a jde podél elektronového transportního řetězce.
Jakou roli hrají elektrony chlorofylu ve fotosyntéze?
Odpovědět
Elektrony chlorofylu, excitované slunečním zářením, procházejí elektronovými transportními řetězci a odevzdávají svou energii tvorbě ATP a NADP-H2.
V jaké fázi fotosyntézy vzniká volný kyslík?
Odpovědět
Ve fázi světla, při fotolýze vody.
V jaké fázi fotosyntézy dochází k syntéze ATP?
Odpovědět
světelná fáze.
Co je zdrojem kyslíku při fotosyntéze?
Odpovědět
Voda (při fotolýze vody se uvolňuje kyslík).
Rychlost fotosyntézy závisí na limitujících (limitujících) faktorech, mezi které patří světlo, koncentrace oxidu uhličitého, teplota. Proč jsou tyto faktory limitující pro fotosyntézní reakce?
Odpovědět
Světlo je nezbytné pro excitaci chlorofylu, dodává energii pro proces fotosyntézy. Oxid uhličitý je potřebný v temné fázi fotosyntézy, z něj se syntetizuje glukóza. Změna teploty vede k denaturaci enzymů, zpomalují se reakce fotosyntézy.
Při jakých metabolických reakcích v rostlinách je oxid uhličitý výchozí látkou pro syntézu sacharidů?
Odpovědět
v reakcích fotosyntézy.
V listech rostlin intenzivně probíhá proces fotosyntézy. Vyskytuje se ve zralých a nezralých plodech? Vysvětlete odpověď.
Odpovědět
Fotosyntéza probíhá v zelených částech rostlin vystavených světlu. Ve slupce zelených plodů tedy dochází k fotosyntéze. Uvnitř plodů a ve slupce zralých (ne zelených) plodů nedochází k fotosyntéze.
Mnoho z nás se tak či onak setkalo se solárními články. Někdo používal nebo používá solární panely k výrobě elektřiny pro domácí účely, někdo používá malý solární panel k nabíjení svého oblíbeného gadgetu doma. polních podmínkách, a někdo jistě viděl malý solární článek na mikrokalkulátoru. Někteří měli dokonce to štěstí, že je navštívili.
Přemýšleli jste ale někdy o tom, jak probíhá proces přeměny sluneční energie na elektrickou? Jaký fyzikální jev je základem fungování všech těchto solárních článků? Vraťme se k fyzice a podrobně pochopme proces generování.
Od samého počátku je zřejmé, že zdrojem energie je zde sluneční světlo, respektive, vědecky řečeno, získává se díky fotonům slunečního záření. Tyto fotony si lze představit jako proud elementárních částic kontinuálně se pohybujících od Slunce, z nichž každá má energii, a proto celý světelný tok nese nějaký druh energie.
Z každého čtverečního metru povrchu Slunce je nepřetržitě vyzařováno 63 MW energie ve formě záření! Maximální intenzita tohoto záření spadá do rozsahu viditelného spektra -.
Vědci tedy zjistili, že hustota energie toku slunečního světla ve vzdálenosti od Slunce k Zemi 149 600 000 kilometrů, po průchodu atmosférou a po dosažení povrchu naší planety, je v průměru přibližně 900 wattů na metr čtvereční.
Zde můžete tuto energii vzít a pokusit se z ní získat elektřinu, tedy přeměnit energii slunečního světelného toku na energii pohybujících se nabitých částic, jinými slovy na.

K přeměně světla na elektřinu potřebujeme fotovoltaický konvertor. Takové měniče jsou velmi běžné, nacházejí se ve volném prodeji, jedná se o tzv. solární články - fotoelektrické měniče ve formě desek vyřezaných z křemíku.
Nejlepší jsou monokrystalické, mají účinnost kolem 18%, to znamená, že pokud má tok fotonů ze slunce hustotu energie 900 W / m2, můžete počítat se 160 W elektřiny na metr čtvereční baterie sestavené z takových článků.
Funguje zde jev zvaný „fotoelektrický efekt“. Fotoelektrický jev nebo fotoelektrický jev- jedná se o jev emise elektronů látkou (jev vytahování elektronů z atomů látky) pod vlivem světla nebo jiného elektromagnetického záření.
Již v roce 1900 Max Planck, otec kvantové fyziky, navrhl, že světlo je vyzařováno a absorbováno v oddělených částech neboli kvantech, které později, konkrétně v roce 1926, chemik Gilbert Lewis nazval „fotony“.

Každý foton má energii, kterou lze určit vzorcem E = hv - Planckova konstanta krát frekvence záření.
V souladu s myšlenkou Maxe Plancka se fenomén objevený v roce 1887 Hertzem a poté důkladně prostudovaný v letech 1888 až 1890 Stoletovem stal vysvětlitelným. Alexander Stoletov experimentálně studoval fotoelektrický jev a stanovil tři zákony fotoelektrického jevu (Stoletovovy zákony):
Při konstantním spektrálním složení elektromagnetického záření dopadajícího na fotokatodu je saturační fotoproud úměrný energetickému osvětlení katody (jinak: počet fotoelektronů vyražených z katody za 1 s je přímo úměrný intenzitě záření).
Maximální počáteční rychlost fotoelektronů nezávisí na intenzitě dopadajícího světla, ale je určena pouze jeho frekvencí.
Pro každou látku existuje červená mez fotoelektrického jevu, tedy minimální frekvence světla (v závislosti na chemické povaze látky a stavu povrchu), pod kterou je fotoelektrický jev nemožný.
Později, v roce 1905, Einstein objasnil teorii fotoelektrického jevu. Ukáže, jak kvantová teorie světla a zákon zachování a přeměny energie dokonale vysvětlují, co se děje a co je pozorováno. Einstein sepsal rovnici pro fotoelektrický jev, za kterou dostal v roce 1921 Nobelovu cenu:

Pracovní funkce A zde je minimální práce, kterou musí elektron vykonat, aby opustil atom látky. Druhý termín - Kinetická energie elektron po výstupu.
To znamená, že foton je absorbován elektronem atomu, díky čemuž se kinetická energie elektronu v atomu zvyšuje o množství energie absorbovaného fotonu.
Část této energie se spotřebuje na výstup elektronu z atomu, elektron atom opustí a dostane možnost se volně pohybovat. A směrově se pohybující elektrony nejsou nic jiného než elektrický proud nebo fotoproud. V důsledku toho můžeme mluvit o výskytu EMF v látce v důsledku fotoelektrického jevu.

to znamená, Solární baterie funguje díky fotoelektrickému jevu, který v ní působí. Kam ale jdou „vyřazené“ elektrony ve fotoelektrickém měniči? Fotoelektrický konvertor nebo solární článek nebo fotočlánek je tedy fotoelektrický jev v něm nastává nezvykle, jedná se o vnitřní fotoelektrický jev a má dokonce zvláštní název „ventilový fotoelektrický efekt“.
Pod vlivem slunečního záření p-n křižovatka Na polovodiči se objeví fotoelektrický jev a objeví se EMF, ale elektrony neopustí fotočlánek, vše se děje v blokovací vrstvě, kdy elektrony opustí jednu část těla a přesunou se do jeho jiné části.
Křemík v zemské kůře tvoří 30 % její hmotnosti, takže se používá všude. Zvláštnost polovodičů obecně spočívá v tom, že nejsou vodiči ani dielektriky, jejich vodivost závisí na koncentraci nečistot, na teplotě a na vystavení záření.
Pásmová mezera v polovodiči je několik elektronvoltů, a to je pouze energetický rozdíl mezi horní úrovní valenčního pásma atomů, ze kterého elektrony unikají, a spodní úrovní vodivostního pásma. Křemík má zakázané pásmo 1,12 eV – přesně to, co je potřeba k absorpci slunečního záření.

Takže přechod p-n. Dopované křemíkové vrstvy ve fotočlánku tvoří p-n přechod. Zde se získá energetická bariéra pro elektrony, ty opustí valenční pásmo a pohybují se pouze jedním směrem, díry se pohybují opačným směrem. Takto se získává proud v solárním článku, tedy probíhá výroba elektřiny ze slunečního záření.
P-n přechod, vystavený působení fotonů, nedovoluje nosičům náboje - elektronům a dírám - pohybovat se jiným směrem než pouze jedním směrem, oddělují se a končí na opačných stranách bariéry. A tím, že je fotovoltaický měnič připojen k zátěžovému obvodu přes horní a spodní elektrodu, vytvoří se ve vnějším obvodu vystavený slunečnímu záření.
souhrn další prezentace"Problematika jednotné státní zkoušky z biologie 2013" - Genotyp organismu. symbiotický vztah. Molekula proteinu. Základní chyby. Tabákový kouř. Podobnosti a rozdíly mezi mutační a kombinační variabilitou. Nukleotid. Kolik buněk se tvoří v důsledku meiózy. Molekuly bílkovin. Krevní skupina. Downův syndrom. Triplet nukleotidů. Etapy energetického metabolismu. Biologická konzultace. krevní destičky. Variabilita kombinací. Schopnost chemo-autotrofní výživy.
"Nemoci vylučovacího systému" - Cystitida. Uretritida. diabetická nifropatie. Nefrogenní anémie. Urolitiáza onemocnění. Pyelonefritida. Hydronefróza. Polycystické onemocnění ledvin. Renální kolika. Amyloidóza ledvin. Akutní onemocnění vylučovacího systému. Prostatitida.
"Období paleogenu" - Podnebí. Podnebí bylo dokonce tropické. Cenozoická éra. Kostnatá ryba. oligocén. Počátek paleogénu. Svět zvířat. Diatryma. paleogén. Vějířoví bezzubí ptáci. svrchní eocén. Paleogenní období.
"Otázky jednotné státní zkoušky z biologie" - rozkvět třídy Bird. Jaký druh rozmnožování je asexuální. Jaká struktura chloroplastu obsahuje enzymy. Vytvořte korespondenci mezi znakem a orgánem. somatické mutace. vzhled tkání. Vnímání podráždění. U obratlovců se orgán sluchu v procesu evoluce změnil. Rozbor výsledků zkoušky z biologie. Placentace. Vznik plic. Struktura oka. Kolik autozomů je v jádrech somatických buněk.
"Pravidla zdravého stravování" - Organizace jídelen plného cyklu. Realizace programu. Kuře. Výsledky realizace programu. Protein. Správná výživa je způsob života. Zdravé jídlo. Správná výživa. Výživové teorie. Školní stravování. Klasická teorie vyvážené výživy. Komplexní reorganizace systému školního stravování. Diety. Cíle a cíle programu. Existují 2 možnosti, jak sestavit školní jídelní plán.
"Výroba mléčných výrobků" - Studium kvality mléka. Konstrukce. živočišný průmysl. Zpráva z turné. Stanovení sacharidů v mléce. Staritsky máslo a sýr rostlina. Věta. Práce kritiků a analytiků. Vyrobené mléko. Příspěvek velkého vědce. Příspěvek Dmitrije Ivanoviče Mendělejeva k rozvoji mlékárenského průmyslu. Skutečný vědec. Objednat. Vývoj sýra. Nápady. Definice tuku. Vlastnosti základní části mléko.